Jerry Bergman
Z CRSQ ročník 46, č.4, 2010, z článku The Pleiotropy, Problem for Evolution přeložil Pavel Kábrt – 04/2010.
Ve zkratce
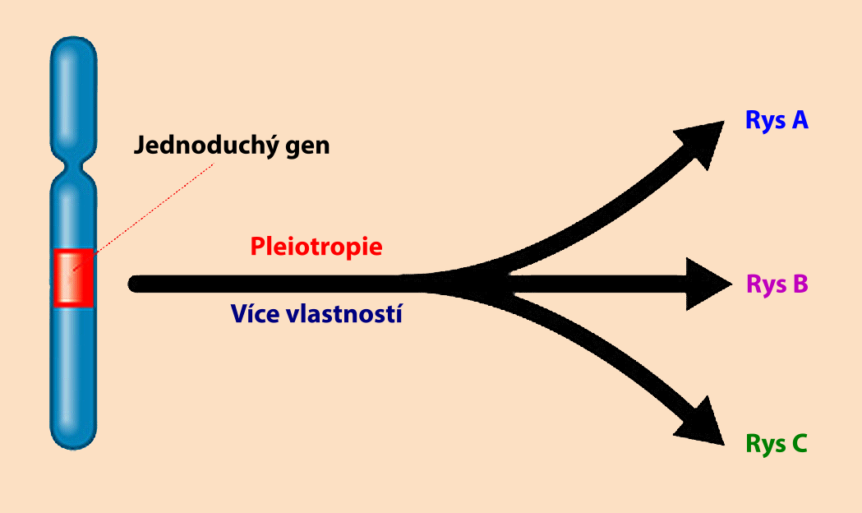
Pleiotropie je jev, při kterém v důsledku propojeného genetického systému jediný gen ovlivňuje mnoho různých biologických systémů, pozitivně i negativně. Dochází k ní proto, že vícečetný fenotypový účinek vzniká z každého exprimovaného genu. V této studii je ukázán nepříznivý účinek pleiotropie na přírodní výběr. V závěru z toho vyplývá, že pleiotropie vytváří velký problém pro evoluční teorii, protože nahromaděné mutace, i ty příznivé, mají často neočekávané negativní účinky.
Úvod
Pleiotropie – z pleio (mnoho) a tropo (změny) – je definována jako situace, ve které jediná genetická proměnná zodpovídá za celou řadu rozličných a často vzájemně nesouvisejících fenotypových účinků (King a Stansfield, 1997, str. 264). Geny nepracují nikdy samostatně, ale jsou součástí vysoce propojeného biologického systému (Wagner et al., 2008). I když je tedy mutace příznivá pro jeden znak, stejně se často projeví i negativní účinky, které vytvoří to, čemu se říká „cena za zdatnost“. U lidí je důsledek zřejmý, kdy jediná mutace způsobí chorobu, která vyprodukuje mnoho vzájemně nesouvisejících symptomů (Dudley et al., 2005).
Nejlépe známým příkladem pleiotropie jsou mnohočetné účinky, které povstávají z porušení jediného genu u srpkovité anemie, nemoci pocházející z jediné mutace v hemoglobinovém genu. Vedle anemie se projeví další komplikace: vředy na nohou, potíže s kostmi, srážení krve, anoxie /nedostatečné prokysličování, poz. př./ vznikající z abnormálních krevních buněk neschopných správného proudění, porušení sleziny, mrtvice a hemolýza /rozpad červených krvinek, erytrocytů, poz. př./. Bakterie také často využívají určitý typ pleiotropie jako součást svých adaptabilních schopností (Anderson a Purdom, 2008).
Pleiotropie, mutace a evoluce
Úskalí pleiotropie musí být důkladně prověřeno, protože mutace se považují za hlavní zdroj genetické variability, ze kterých vybírá selekce pro evoluční změnu. Pleiotropie je „opakovaně pozorována“ v přírodě a má zcela „zřejmé evoluční nevýhody pro organizmus, včetně omezené míry adaptace pro některé znaky v důsledku selekce jiných“ (Dudley et al., 2005, str. E8). Ta nevýhoda pro evoluci mutacemi je tato: Jestliže je určitý fenotyp /vlastnost organizmu, poz. př./ výsledkem mutace, která způsobí ztrátu nebo změnu jediné funkce, může v důsledku toho dojít ke ztrátě jiných důležitých funkcí, protože se poruší část větvících se cest.
Tato překážka evolučnímu pokroku se nazývá „cena za složitost“ (Orr, 2000, str. 13). Další překážkou je, že ztráta funkce u jednoho genu může nepříznivě ovlivnit několik fenotypových znaků, a obtíže mohou nastat z toho, když je protein kódovaný jedním genem nutný pro několik funkcí (viz Dudley et al., 2005).
Pleiotropie je důsledkem té skutečnosti, že většina genů eukaryotických buněk je řízena deseti i více regulačními proteiny, a každý gen je střídavě řízen více jak tuctem různých vazebních regulačních proteinových míst. Tato místa, odkud jsou geny řízeny, jsou obvykle v těsné nadřazenosti vůči regulovaným genům, přesto však mohou být umístěna v nějaké vzdálenosti, dokonce na jiném chromozomu. Z tohoto důvodu může mít jedna mutace mnohočetné fenotypové důsledky, často v různých orgánech a dokonce v různých orgánových systémech (Hodgkin, 1998).
Samostatnou skupinou genů jsou geny homeotické; ty všechny obsahují sekvence nazývané homeobox. Ty produkují tzv. hox proteiny. Hox proteiny jsou hlavními řídícími jednotkami (master regulators) stavebních programů u koordinovaných konstrukcí celých struktur, jako jsou křídla nebo končetiny. Mutace v hox genech mohou způsobit celou sérii velkých fenotypových změn, jako je třeba vznik další sady nefunkčních křídel. Často mutace v hox genech vyprodukují kaskádu změn, které nakonec vyústí ve smrt organizmu.
Podobně jako výstavba moderního kancelářského komplexu vyžaduje koordinaci stovek či tisíců lidí, analogicky totéž vyžadují konstrukce buněk, křídel a orgánů jako jsou plíce, srdce či ledviny. Budova nebude správně fungovat, pokud mechanické, elektrické a konstrukční prvky nejsou správně vyprojektovány, nebo jsou nesprávně sestaveny a/nebo nevhodně propojeny. Stejně tak, jsou-li části orgánu nebo struktury nesprávně složeny nebo mají projektové chyby, organizmus nebude životaschopný.
Typy pleiotropie
Geny jsou klasifikovány do dvou základních typů vzhledem k jejich pleiotropním účinkům: vysoce pleiotropní geny nazývané pleiotropní geny s velkým účinkem (major geny) a nepleiotropní geny či omezené pleiotropní geny, nazývané pleiotropní geny s malým účinkem (minor geny), které mají nepatrný účinek, protože asi fungují nezávisle (Albert et. al., 2007). Tato klasifikace je však omezená, protože přestože některé geny jsou „major“ a některé „minor“, mnohé vykazují pleiotropní účinek na fenotyp, ze zcela jiných důvodů.
Hodgkin (1998) navrhl klasifikační systém rozdělující pleiotropní účinek na sedm různých typů dle příčiny. Tento klasifikační systém dokumentuje, jak silně jsou geny na fenotypové úrovni propojené i to, jakými různými cestami se toto propojení uskutečňuje. Jeho klasifikační systém je tento:
1. Artefaktuální /nepřirozená/ pleiotropie, při které sousední avšak funkčně nezávislé geny jsou ovlivněny jedinou mutací, např. když jsou dva geny umístěny na chromozomu naproti sobě a mutace jednoho ovlivní i druhý. Hodgkin to uvádí tak, že organizmy s „kompaktními, genově zhuštěnými genomy, budou zvláště citlivé na artefaktuální pleiotropii“ (1998, str. 502). Toto pozorování ukazuje, že tato pleiotropie může být překážkou spíše v jednodušších a primitivnějších organizmech. Příkladem je claret-nondisjunktní mutace u Drosophily, způsobující jak abnormality v barvě očí, tak i meiotickou nondisjunkci /chyba v rozcházení chromozómů k opačným pólům buňky při meióze, když se homologické chromozómy od sebe neoddělí – poz. př./.
2. Sekundární pleiotropie nebo „příbuzenecká pleiotropie“ je zapříčiněna jedinou mutací, způsobující biochemické změny ovlivňující mnoho strukturálních změn. Příkladem je mutace způsobující fenylketonurii, která je důsledkem defektního enzymu v játrech (fenylalaninhydroxylázy). Výsledkem je nedostatečná myelinizace axonů /vytváření bílé lipidové hmoty v mozku a míše – poz. př./. Vede k nesčetným zdravotním potížím včetně mentální retardace (Hodgkin, 1998, str. 502). Sekundární pleiotropie je zvláště běžná u složitých organizmů s dlouhým životem, a tak v důsledku představuje velký problém pro evoluci „vyšších“ organizmů.
3. Adoptivní nebo exaptační pleiotropie vzniká v situaci, kdy je jeden genový produkt užíván pro mnoho různých biochemických reakcí v různých tkáních. Příklad, který Hodgkin uvádí, je krystalinový protein, nejen hojný v očních čočkách, ale využívaný též pro svoji strukturní úlohu v jiných tkáních, jako je např. hladké svalstvo.
4. Parsimoniální /úsporná/ pleiotropie se vyskytuje tehdy, když je jeden enzym využíván ke katalýze stejné chemické reakce v mnoha různých tkáních a orgánových systémech, nebo je používán v různých biochemických cestách. Příkladem může být skutečnost, kdy ty samé enzymy jsou využívány v různých větvích biochemických cest při syntéze isoleucinu a valinu.
5. Oportunistická pleiotropie je událost, při které jeden regulační protein vykonává důležitou roli u jiné buňky nebo jiných typů tkání jako vedlejší funkci k jeho hlavním funkcím. Příklad, který použil Hodgkin, jsou řídící prvky sisB a runt na chromosomu X, které způsobují problémy v raných fázích vývoje, ale jejichž geny jsou rovněž využívány v pozdějších fázích růstu, např. v sekundárním sexuálním vývoji.
6. Kombinační pleiotropie je případ produktu jednoho genu reagujícího s různými proteiny v různých typech buněk a typech různých cest, pro které je užíván, což vede k odlišné variabilitě. Existuje velké množství příkladů, včetně faktorů většiny transkripce, způsobujících velmi různou biochemickou aktivitu v závislosti na tom, kde k reakci s genomem dojde. Výsledkem pak je, že mutace ovlivňující tento protein „mají mnohočetné a často velmi různorodé účinky na širokou škálu tkání“ (Hodgkin, 1998, str. 503).
7. Sjednocující pleiotropie představuje jev, kdy jeden gen anebo celá skupina sousedících genů kóduje rozličné proteiny, které mají společné nebo příbuzné biologické funkce. Příklady zahrnují různé strukturální komponenty, vazební domény a enzymy. Výsledkem je, že genové mutace v této kategorii „mají složité fyziologické důsledky, což může být obtížně vysvětlitelné, pokud není porozuměno biologickým příčinám ležícím v základech“ (Hodgkin, 1998, str. 503).
Ve všech těchto sedmi typech pleiotropie mohou mutace (i kdyby byly prospěšné) způsobit negativní nebo smrtelné důsledky na mnoha místech. Tento výčet ilustruje mnoho způsobů, kterými jsou všechny systémy spojeny. Také to ukazuje, jak jedna genetická změna může způsobit nepříznivé modifikace v přímo zasažené soustavě, nebo dokonce i v pozdějších vývojových stadiích. To je zásadní obtíž pro evoluci, protože jakákoliv mutace, i ta příznivá, bude mít velmi pravděpodobně nějaké či mnohé negativní důsledky.
Antagonistická pleiotropie
K antagonistické pleiotropii dochází tak, že jeden gen produkuje mnoho navzájem si konkurujících účinků. Příznivé účinky jsou pak vykompenzovány zhoubnými důsledky vyprodukovanými tím samým genem. Například některé geny mohou zvýšit zdatnost (fitness), když je člověk mladý, ale v pozdějším věku ji snížit. Máme na mysli gen pro hormonální testosteron, který zvyšuje reprodukční zdatnost u adolescentů, ale v pozdějším věku může způsobit rakovinu prostaty.
Funkce pleiotropie v biologii
Pleiotropie je nutná pro koordinaci tělesného růstu a tělesný rozvoj. Například normální zdravá malá žena má možnost dosáhnout stejných tělesných proporcí svého těla, jako jsou končetiny, srdce, plíce či jiné vnitřní orgány, jako má tuto možnost normální zdravý vysoký muž. Vysoký muž má delší končetiny než malá žena, ale každá končetina je vázána na ten samý poměr u každé osoby. Pleiotropie je rovněž hlavním prostředkem koordinace různých tělesných funkcí, jako je třeba metabolismus.
Jeden ze způsobů, jak geny ovlivňují fenotypovou korelaci, je to, že geny řídí množství sekretovaných hormonů, a hormony často ovlivňují mnohé oddělené funkce. Příkladem může být růstový hormon sekretovaný hypofýzou, který nezvyšuje pouze výšku a rozměry rukou a nohou, ale řídí veškeré rozměry těla. Mnoho genů má dobře známé pleiotropní funkce, včetně důležitého genu nazývaného p53 a všech genů „heat-shock“ (geny pro proteiny teplotního šoku), které jsou součástí genetického systému reagujícího na teplotní fluktuace (např. Meyers, 1995, str. 281-285). Odezva na teplotní šok aktivuje řadu genů k ochraně před teplotním poškozením. Tato složitá odpověď vůči nebezpečí z teplotního poškození je jak rychlá, tak i vratná.
Jiným příkladem důsledků pleiotropie je cytokinová síť. Cytokiny jsou polypeptidy podobné hormonům, které jsou občasně produkovány mnoha různými buňkami. Obvykle působí lokálně tak, že změní buněčnou fyziologii. Navážou se na buněčné povrchové receptory, aby aktivovaly určité geny. Účinky cytokinů jsou redundantní (tím se myslí to, že různé cytokiny mohou mít ten samý účinek), a tak mohou mít různý dopad na tu samou buňku v rozličných situacích.
Tento fakt opět ilustruje, že jednotlivé mutace, byť by byly příznivé, přesto mají široké spektrum výsledků, z nichž některé jsou naprosto negativní na další systémy. Jediný cytokin může mít různé účinky na různé buňky a různé cytokiny mohou navzájem reagovat buď v součinnosti, nebo proti sobě, nebo jako doplněk (Meyers, 1995). Jelikož původ nové genetické informace, ze které evoluce vybírá, jsou mutace, pleiotropie demonstruje, že pozitivní mutace obvykle zvyšují „cenu za složitost“ tím, že přinášejí více škody než užitku.
Příklady pleiotropie z medicíny
Všechny léky ovlivňují biochemii těla a všechny léky mají vedlejší účinky následkem pleiotropie. Důvodem je to, že všechny léky buď zeslabují anebo aktivují určité aspekty biochemie těla. Mají medikamentózní dopad a vliv na celý propojený biochemický systém. Běžné společné vedlejší účinky mnoha léků jsou únava, nevolnost, suchá ústa, snížená pozornost, žaludeční problémy a alergie. Aspirin zasahuje do biochemické cesty prostaglandinu /tj. látka podobná hormonu – poz. př./, a tím nejen blokuje bolest, ale také schopnost těla chránit žaludek od rozkladných účinků žaludeční kyseliny HCl.
Mutace, která způsobí nemoc a zničí jeden gen, často vyústí v mnoho jiných, časově nesouvisejících symptomů. Mutace, která zničí gen fenylalaninhydroxylázy, produkující enzym pro přeměnu aminokyseliny fenylalanin na aminokyselinu tyrosin, může zvýšit koncentrace fenylalaninu na toxické úrovně. Tato metabolická nemoc, která zde už byla zmíněna, známá jako fenylketonurie (PKU), není-li léčena, způsobuje celou plejádu symptomů včetně mentální retardace, omezení růstu vlasů, modré oči a v důsledku omezené pigmentace také světlost kůže (Hodgkin, 1998).
Všechny tyto příklady ukazují na velký rozsah biochemického propojení lidské fyziologie, a dále na to, jak pokusy něco změnit pro blaho pacienta často vyústí v nechtěné účinky. Stejně tak i užitečná mutace, i kdyby byla velice prospěšná, může způsobit četné nechtěné vedlejší účinky. Cena za složitost může být vysoká, a z tohoto důvodu evoluce pomocí hromadění mutací je velmi problematická. Spíše jsou tyto příklady pleiotropie argumentem pro stvoření ex nihilo.
Příklady pleiotropie z oblasti chovu zvířat
Pleiotropie je význačnou překážkou úspěšného šlechtění zvířat. Negativní dopady, které jsou výsledkem živočišného šlechtění, ilustrují, proč je evoluce hromaděním mutací nerealistickým mechanizmem. Když se zvířata kříží, aby se vyšlechtil jeden kýžený znak, téměř vždy vzniknou i neblahé důsledky (Grandin a Johnson, 2005). Šlechtitelé kuřat vybírají slepice a kohouty, kteří rychle rostou, jsou tlustí a mají dobrou svalovinu. Grandin a Johnson (2005) prokazují, že zisk jednoho takového znaku pravidelně vede ke ztrátě znaků jiných. Například rychle rostoucí slepice a kohouti obvykle skončí na problému plodnosti, což je vedlejší účinek pozorovaný i na Belgickém modrobílém plemenu dobytka a některých dalších zvířatech.
Když byl znak rychlého růstu u kuřat vyselektován, výsledná kuřata měla slabá srdce. Smrt v důsledku srdečního selhání pak byla častější. Snahy vypěstovat kuřata s větším hrudníkem pak vyústily v deformované, křivé a nateklé nohy. Ve snaze vyřešit tento problém chovatelé usilovali vytvořit kuřata jak s většími hrudníky tak i silnějšími srdci. Nakonec vyprodukovali to, co vypadalo jako kuře jejich snů – silná kuřata s většími hrudníky, silnýma nohama a silnými srdci – ale krátce poté zjistili vážné problémy v jejich chování. Tím byla tendence kohoutů zabíjet slepice, částečně v důsledku toho, že šlechtění vedlo k zániku tance, při kterém se kohout dvoří slepicím. Bez tohoto tance slepice sexuálně s kohouty nespolupracovaly. Problém vyšlechtění jednoho znaku při ztrátě jiných znaků je všeobecný. Grandin a Johnson (2005, str. 72) dochází k závěru, že tato nepříznivá okolnost nastává „pokaždé, kdy chovatelé přeženou selekci jednoho znaku. Výsledkem je zdeformovaná evoluce.“
Přesto Grandin a Johnson (2005, str. 72-73) tvrdí, že to, co nedokáže umělá selekce, dokáže ta přírodní. V oddílu nadepsaném „Selekční tlak“ diskutují evoluci barevného vidění u primátů. Tvrdí, že přírodní výběr vyprodukoval barevné vidění u primátů bez inteligence. Z toho plyne, že inteligentní chovatelé obvykle narazí na genetický problém, když se snaží vybrat určité znaky, ale slepé síly přírody tento úkol zvládnou na jedničku. Akorát pro svoje tvrzení neposkytují žádný důkaz.
Negativní následky selekce lze typicky obejít výběrem jen malých vylepšení; tedy například místo největšího možného kuřete vybrat prostě kuřata jen o něco větší a udělat kompromis s jinými znaky, aniž by se pak šlechtitelé museli kompromitovat s nepřekonatelnými a vážnými problémy. Jak je dobře známo, křížení vždy přináší limity, a jak na tyto meze narazíme, vzniknou komplikace v důsledku pleiotropie. A to vše tedy také ukazuje, proč je makroevoluce pomocí hromadění mutací velice nepravděpodobná – jeden krok kupředu trvale způsobí několik kroků zpět následkem pleiotropie.
Epistáze
(genová nadřazenost – pozn. př.)
Koncept pleiotropie je spojen s konceptem epistáze. Oba koncepty se týkají interakcí, které ovlivňují fenotypové znaky. Epistáze odkazuje ke složitým interakcím, které existují mezi různými geny, často když jeden gen je modifikován jinými geny, které nazýváme modifikátory. Synergická epistáze je pozitivní interakce, která vyústí v mohutnější expresi znaku; antagonistická epistáze je negativní interakce vedoucí k nižší expresi znaku.
Epistáze se často váže k normálnímu genetickému řízení a regulaci, zatímco pleiotropie se vztahuje jak k normální, tak i abnormální genové interakci. Ale oba systémy mohou vytvořit vážné problémy darwinistické evoluci založené na kumulaci mutací. Je-li v epistatickém systému změněn jeden gen, může to ovlivnit jiné geny. Evolucionisté argumentují tím, že synergická epistáze dává přírodnímu výběru mocný nástroj k odstranění škodlivých mutací a zabraňuje jejich kumulaci. Uvažují tak, že pokud mutace v jednom genu ovlivní fungování jiných genů, pak mutace, které nejsou letální (smrtící) se mohou stát letální nebo nepříznivě ovlivnit přežití organizmu. A tím organizmus vlastnící mutaci bude pravděpodobněji negativně vyselektován přírodním výběrem, a tím se celkový objem mutací v organizmu sníží.
Zatímco by alespoň teoreticky tento efekt mohl fungovat, může způsobit mnoho normálních neletálních neutrálních mutací (včetně těch potenciálně prospěšných), které se stanou letálními (smrtelnými) nebo negativně vyselektovanými v důsledku epistatických interakcí. Následně pak synergický účinek odstraní nejen škodlivé mutace, ale i většinu těch potenciálně prospěšných. Čistým výsledkem bude odstranění jak negativních tak i pozitivních mutací, tedy konzervační účinek, který vzdoruje genetickým změnám. Za zvýšení prospěšných mutací a jejich zafixování v populaci bude zvýšena cena za selekci (ReMine, 2006). Je nepravděpodobné, že většina populací bude schopna tuto zvýšenou cenu zaplatit.
Molekulární mechanizmus pleiotropie
Jeden z důležitých důvodů pleiotropie je, že molekuly musí v živém organizmu neustále interagovat (reagovat), jedna s druhou. Aby fungovaly, musí se vázat na některé další molekuly, ale ne na jiné. Nejčastěji molekuly interagují s mnoha jinými molekulami v buňce v rámci tzv. „hlučného biochemického prostředí“ (Savir a Tlusty, 2007, str. e468). Mutace často mění konformitu (tvar) a dokonce vazební vlastnosti molekul, což může ovlivnit jejich interakce s dalšími molekulami.
Tento důsledek je důležitý, protože „prakticky všechny biologické systémy jsou odkázány na schopnost biomolekul specializovaně rozpoznat jedna druhou“, a změna může způsobit, že molekula nerozezná tu, kterou rozeznat má, a následně bude reagovat s tou, se kterou reagovat nemá, což obé způsobuje problémy (Savir a Tlusty, 2007, str. e468). K některým běžným příkladům patří rozpoznávací schopnost protilákta-antigen, navázání se regulačních proteinů na DNA a enzym-substrát interakce nutné pro fyziologické chemické reakce.
Překrývající se geny
Překrývající se geny se vyskytují tam, kde jeden či více genů existuje uvnitř jiného genu, nebo se dva geny překryjí tak, že se gen stane součástí genu sousedního. Existují důkazy, že až 12 či více sekvencí se překrývá v jednom genu. Změna jednoho nukleotidu tak může ovlivnit mnoho informačních systémů, což povede k tomu, že téměř všechny mutace budou mít pleiotropní účinky (Trifonov, 1989; 1997). Nedávný projekt ENCODE prokázal, že typický nukleotid je částí mnohonásobně se překrývajícího sdělení, a to ukazuje, že většina mutací bude mít pleiotropní účinek (Kapranov et. al., 2007). Sanford představil koncept „polyfunkční DNA“, což znamená, že mnoho funkčních jednotek DNA je „mnohovazebných“. Z toho plyne, že mutace, která vylepší jedno sdělení (zprávu, informaci) často zničí ta sdělení, která to první překrývají (Sanford, 2008).
Jak moc je pleiotropie častá?
Studie mnoha životních forem, včetně kvasinek, dokládají, že existuje „velký počet pleiotropních genů“ (Dudley et al., 2005). Hodně výzkumných prací ukazuje, že většina, a snad všechny geny, mají pleiotropní účinky. Přinejmenším je dosud jasné, že většina genů je má. Například samotné oblasti promotorů /kde se zahajuje transkripce – poz. př./ jsou řízeny od deseti až do desítek míst s transkripčními faktory (Stone a Wray, 2001). Jedna taková studie, která se zabývala množstvím znaků ovlivněných jednotlivou mutací, použila analýzu QTL (quantitative trait loci – jde o zjištění lokusu, který výrazně ovlivňuje variabilitu ve zkoumaném znaku – poz. př.) pro znaky na kostře myší. Zjistila, že polovina QTL ovlivnila až šest znaků a jedna sada QTL ovlivnila od 25 až ke 30 různých znaků (Wagner et al., 2008, str. 471).
Jiná QTL studie se zaměřila na počet genů s velkým a malým pleiotropním účinkem (major a minor geny) na tvar těla koljušky /asi 5-10 cm velká rybka – poz. př./. Výsledek ukázal, že asi polovina genů měla minoritní pleiotropní účinek a několik mělo „velké a zřejmě široce rozšířené účinky“ (Albert et al., 2007, str. 76). Je třeba více studií k určení úrovně pleiotropních účinků dalších genů. Pravděpodobný účinek je mnohem větší než tato studie ukazuje.
Hox geny a další transkripční faktory jsou známé tím, že ovlivňují a regulují morfologii při vývoji těla (Coyne, 2005). Hodgkin (1998, str. 501) tvrdí, že pleiotropie „bude spíše u vyšších organizmů pravidlem než výjimkou.“ Ukázal, že nedávné důkazy podporují tento závěr, což ukazuje jeho důležitost u vyšších organizmů. Jaroslav Flegr navrhl, že pleiotropie omezuje plasticitu druhů (Hall, 2009). Vysoká plasticita životních forem, třeba psů, umožňuje produkci mnoha morfologických variací, a plastické druhy mají větší podíl svých znaků kódovaných jediným genem. Vykazují tak méně interakcí typu gen-gen, zatímco zamrzlé druhy mají větší část svých znaků kódovaných větším počtem genů a vykazují zvýšené genetické interakce (Hall, 2009).
Shrnutí
Pleiotropie nejen vytváří velkou genetickou bariéru jak pro mikro a makro evoluci, ale dokonce též staví limity pro šlechtění zvířat a rostlin v důsledku biochemické provázanosti existující v buňkách, tkáních, orgánech a orgánových systémech. Tato skutečnost je velmi dobře známa rostlinným a živočišným šlechtitelům, stejně jako těm, kteří sní o čistokrevně vyšlechtěných psech, koních a jiných zvířatech. Jak to připustil Hodgkin (1998, str. 501), „Ve složitých eukaryotech může vést pleiotropie k velkým překážkám pro případné mutační cesty“, umožňující evoluci. Jak již bylo řečeno, důkazy pro pleiotropii favorizují speciální stvoření (Tinkle, 1075).
Poděkování
Moje poděkování si zaslouží Clifford Lillo, Nancy O´Sullivan, PhD, Ted Siek, PhD, Tom Hennigan, George Howe, PhD a John UpChurch za prohlédnutí prvního konceptu této práce.
Odkazy na literaturu
Albert, A.Y.K., S. Sawaya, T. Vines, A.K. Knecht, C.T. Miller, B.R. Summers, S. Balabhadra, D. Kinksley, and D. Sclucter. 2007. The genetics of adaptive shape shift in stickleback: pleiotropy and effective size. Evolution. 62-1:76–85.
Anderson, K.L., and G. Purdom. 2008. A creationist perspective of beneficial mutations in bacteria. In A.A. Snelling
(editor), Preceedings of the Sixth International Conference on Creaitonism, pp. 73–86. Creation Science Fellowship
(Pittsburg, PA) and Institute for Creation Research (Dallas, TX).
Coyne, J. 2005. Switching on evolution. Nature 435:1029–1030.
Dudley, A.M., D.M. Janse, A. Tanay, R. Shamir, and G.M. Church. 2005. A global view of pleiotropy and phenotypically
derived gene function in yeast. Molecular Systems Biology doi:10.1038/ msb4100004–E1.
Grandin, T., and C. Johnson. 2005. Animals in Translation. Scribner, New York, NY.
Hall, B.K. 2009. Review of Frozen Evolution: Or That’s Not the Way It is, Mr. Darwin, edited by Jaroslav Flegr. Evolution & Development 11(1):126–129.
Hodgkin, J. 1998. Seven types of pleiotropy. International Journal of Developmental Biology. 42:501–505
Kapranov, P., A.T. Willingham, and T.R.Gingeraw. 2007. Genome-wide transcription and the implications for genome
organization. Nature Reviews Genetics 8:413–423.
King, R., and W. Stansfield. 1997. A Dictionary of Genetics. Oxford University Press, New York, NY.
Meyers, R. 1995. Molecular Biology and Biotechnology. VCH Publishers, New York, NY.
Orr, H.A. 2000. Adaptation and the cost of complexity. International Journal of Organic Evolution 54:13–20.
ReMine, W. 2006. More precise calculations of the cost of substitution. Creation Research Society Quarterly 43:111–120.
Sanford, J.C. 2008. Genetic Entropy and the Mystery of the Genome. FMS Publications, Waterloo, NY.
Savir, Y., and T. Tlusty. 2007. Conformational proofreading: The impact of conformational changes on the specifi city
of molecular recognition. PLoS One 2: e468.
Stone, J.R., and G.A. Wray. 2001. Rapid evolution of cis-regularity sequences via local point mutations. Molecular Biology
Evolution 18:1764–1770.
Tinkle, William J. 1971. Pleiotropy—extra cotyledons in the tomato. Creation Research Society Quarterly 8:183–185.
Tinkle, William J. 1975. Further research on reduced viability of mutant plants. Creation Research Society Quarterly 12:52.
Trifonov, E.N. 1989. Multiple codes of nucleotide sequences. Bulletin of Mathematical Biology 51:417–432.
Trifonov, E.N. 1997. Genetic sequences as products of compression by inclusive superposition of many codes. Molecular
Biology 31:647–654.
Wagner, G., P. Kenney-Hunt, M. Pavlicev, J.R. Peck, D. Waxman, and J. M. Cheverud. 2008. Pleiotropic scaling of gene effects and the “cost of complexity.” Nature 452:470–472.
Jerry Bergman, PhD:
Biology Department, Northwest State College,
Archbold Ohio 43543, jbergman@northweststate.edu