
Z odborné literatury vybral Pavel Kábrt.
Základní přístup evolucionistů k pravdě je tento: PŘÁNÍ JE OTCEM MYŠLENKY. Pravda se v evolucionizmu neodvozuje od poznaných faktů, nálezů či zákonů vědy, ale od evolucionistické, přes 150 let rozšiřované ideologické myšlenky, že všechno na tomto světě se „v minulosti udělalo samo“. A všechna následná, postupně zjišťovaná fakta, se této myšlence musejí přizpůsobit. Už po mnoho desítek let je hlavní pracovní náplní evolucionistů (umělecky maskované) všívání nepříjemných faktů do lajdácky spíchnutého (a tolikrát záplatovaného) pláště své vetché hypotézy.
1. Úvodem
Všechny přírodovědecké obory (skutečné vědy) jsou pro evoluční fantazie traumatem. Třeba genetika, velký současný problém pro evoluční teorii – jenže i ta je pro sériovou výrobu evolučních mýtů pořád ještě „příliš štědrá“. Ona totiž počítá s již existující, replikující se buňkou včetně funkčního genetického systému. Stejně tak se opírá o „dary z nebes“ evolucionistická argumentace přírodním výběrem – ten také už rovnou předpokládá existenci objektů, schopných předávat určitou vybranou informaci.
Tento článek je o vzniku života. Jeho záměrem je rázně zastavit všechny ústupky evolučním pohádkám. A tak se pojďme podívat na to, co všechno je nutné ke vzniku reprodukčního systému – aby mrtvá hmota sama od sebe ožila, tedy vytvořila buňku, a ta se dále množila.
Původ života je pro evoluční materialisty skutečně velmi bolestné téma. I ten nejjednodušší jednobuněčný organismus je extrémně složitý, zahrnuje velké množství složitých mechanizmů a instrukcí ke své stavbě. A to všechno je navíc uloženo takovým způsobem, že to může být dekódováno jak pro samotnou potřebu organismu, tak pro předání dalšímu potomstvu. I ten nejjednodušší reprodukující se organismus, jaký je možné si představit a který má fungovat, vyžaduje velké množství proteinů, molekulárních zařízení [biologických strojů] a mechanizmus pro zakódování a uložení potřebné informace.1 Je možné, aby se takováto buňka vyvinula z předchozích chemických součástí?
Ve všech známých reprodukujících se organizmech na této zemi je úložištěm biologické informace DNA. Tuto informaci však lze přečíst jen pomocí dekódovacího mechanizmu. Vlastní instrukce pro vytvoření tohoto dekódovacího mechanizmu jsou rovněž uloženy v DNA. Je možné vyluštit tento děsivý problém typu „co bylo dřív, vejce nebo slepice“? A dále – většina těchto procesů používá energii, kterou jim dodává ATP. Ten je vytvářen nanomotorem ATP syntázou. Jenže motor ATP syntáza nemůže vzniknout bez instrukcí v DNA, které zase přečte jen dekódovací mechanizmus používající ATP – jde o trojcestný okruh. Jinak řečeno, jde o problém vajíčko-larva-kobylka (tedy ještě horší než u problému vejce-slepice). Má tento nepochopitelný hlavolam nějaké řešení, nebo nám to říká, že samovolný vznik života, jak jej známe, není možný?
Někteří ve snaze tento problém vyřešit ukazují na existenci jednoho typu molekuly, která by fungovala jako systém pro uchování/zpřístupnění informace a zároveň jako dekódovací mechanizmus. Nejběžnějším kandidátem pro toto řešení bývá RNA. Jak se tedy vyrovná s požadavky na první život?
K tomu je třeba říci, že neřízené chemické procesy, v nějaké hypotetické pravěké chemické polévce, by neměly žádnou šanci dosáhnout ani této minimální složitosti. Předpokládané stavební kameny (aminokyseliny) by nevytvořily potřebně dlouhé molekuly, jaké jsou pro život nutné; naopak, dlouhé molekuly by se rozpadly. Mnoho stavebních kamenů by se vůbec nevytvořilo, nebo by byly tak naředěné a kontaminované, že by se nedaly použít. Byly by to směsi levo-pravotočivých forem, namísto čistě jen jedné či druhé formy, které živé organizmy používají. Jsou tato fakta umíráčkem pro všechny scénáře vzniku života?
K tomu evolucionistické chemické studie, prováděné ve zkumavce nebo simulované na počítači, jsou v nepřijatelné míře ovlivňovány inteligentním badatelem (tedy tím, co u pravěké chemické polévky nebylo k dispozici). Mohou být tedy předkládány jako důkazy vzniku života?
A k dovršení všeho přední badatelé připouštějí, že chemická evoluce je přijímána ne v důsledku faktů a důkazů, ale v důsledku materialistické víry. Jak by tohle všechno bylo použitelné jako nějaký důkaz proti tvrzení Bible, že život stvořila inteligentní Bytost aktem svého mocného působení?
2. Vznik prvního života nelze evolučními procesy vysvětlit
Mezi myšlenkou, že život vznikl chemickými procesy a mezi darwinistickou evolucí existovalo vždy napětí. Jenže většina lidí nemá o tomto sporu ani ponětí. Zdá se, že sám Darwin se potýkal s otázkou vzniku života ve snaze ji sladit se svou teorií. Tiskem zmínil několik opatrných výroků, více poznámek od něj máme v jeho osobních dopisech. Většinou ale jeho písemnosti vykazují celkovou snahu vyhnout se nějakým konečným závěrům. Věděl, že staré spekulace o spontánním plození (samoplození, jako třeba myší či much) vyvrátil Francesco Redi v roce 1668, ale pak by se ještě musel doslova poprat s mocnými argumenty svého současníka, kreacionisty Louise Pasteura, který spontánní plození – také pro mikroorganizmy – naprosto demaskoval v roce 1861,2 jen dva roky po vydání knihy O původu druhů.

Louis Pasteur ve své laboratoři. Foto: Wikipedia.com
V závěrečné kapitole prvního vydání knihy Origin (O původu druhů, 1859) Darwin napsal:
- Analogicky bych měl odvodit, že zřejmě všechny organické bytosti, které kdy žily na této zemi, pocházejí z nějaké prvotní formy, do které byl život nejprve vdechnut.3
Tohle je asi stěží nějaké potvrzení spontánního původu života. Když psal později svému příteli, v roce 1863, litoval, že užil tato slova, ale směr svého uvažování o této věci zachoval:
- Dlouho už lituji, že jsem se chtěl zavděčit veřejnému mínění, a použil jsem výraz z Pentateuchu [biblických pět knih Mojžíšových] pro stvoření, čímž jsem ale ve skutečnosti myslel ´objevil se´ nějakým neznámým procesem…V současnosti je zcela k ničemu přemýšlet o původu života; stejně tak by někdo mohl přemýšlet o původu hmoty.4
Jak zvláštní, že tuto závěrečnou větu vložil do každého dalšího vydání Původu.
- Jest vznešenost v tomto názoru na život s jeho několika mohutnostmi, jež byly původně vdechnuty Stvořitelem v málo tvarů aneb jen v jeden; a že se, zatímco tato oběžnice pokračovala kroužíc dle pevných zákonů všeobecné tíže, z tak prostého počátku vyvinulo a vyvíjí nekonečně těch nejkrásnějších a nejpodivuhodnějších tvarů. [Klapálkův překlad z roku 1914]
Že Darwin věřil v neřízenou evoluci je jasné – není však jasné, zda věřil v evoluci chemickou. Poté se v roce 1871 (to je rok po vydání knihy O původu člověka, ve které poprvé jasně vysvětlil evoluční vztah člověka k nižším formám života) vrhl střemhlav do sporu:
- …jestliže (a Ach! to je velké jestliže!) bychom v nějakém malém teplém jezírku, se všemi přítomnými druhy amonných solí a fosforečnanů, světlem, teplem, elektrickou energií atd. pozorovali, jak se chemicky formuje sloučenina proteinu [sic] připravená procházet stále dalšími složitějšími změnami…5
A tak i Darwin, otec moderní evoluční teorie, kulhal na obě strany mezi dvěma názory na nejdůležitější otázku ze všech – jak vznikl život?
Na tento problém ukázal proslulý filozof Antony Flew (1923-2010), když odpověděl přímo na tvrzení vůdčího ateistického evolucionisty Richarda Dawkinse6 (1941- ):
- Připadá mi, že Richard Dawkins neustále přehlíží fakt, že ve čtrnácté kapitole O původu druhů sám Darwin poukázal na to, že jeho celé tvrzení začalo živou podstatou, která má sama o sobě schopnosti reprodukce. To je takový tvor, jehož evoluci musí teprve evoluční teorie nějak vysvětlit, pokud je sama pravdu zahrnující teorií.
Darwin si byl velmi dobře vědom toho, že toto vysvětlení nepřinesl. A teď mi to připadá tak, že více jak padesátileté objevy z výzkumu DNA poskytly materiály pro nový a nesmírně silný argument ve prospěch designu.7
To je zvláště významné, protože Dr. Flew byl většinu svého života zastáncem ateismu; ale ke zděšení ateistické společnosti tuto víru před svou smrtí opustil.8 Jedním z hlavních důvodů tohoto jeho rozhodnutí je ohromná složitost i té nejjednodušší replikující se buňky.
To samé si uvědomil Theodosius Dobzhansky (1900-1975), jeden z vůdčích evolucionistů 20. století a zapřisáhlý materialista (i přesto, že byl vychováván v ruské ortodoxní víře9). Ve svém komentáři k původu života razantně odmítá teoretiky, kteří se ve svém vysvětlování dovolávají přírodního výběru, protože ten vyžaduje již předem existující život:
- Při četbě další literatury o původu života se obávám, že ne všichni autoři použili termín [přírodní výběr] pečlivě. Přírodní výběr je rozlišující reprodukcí, neustálé trvání organismu. Aby mohl probíhat přírodní výběr, musíte mít sebe reprodukci či sebe replikaci a alespoň dvě rozdílné, sebe replikující jednotky či objekty. … Rád bych vás požádal, docela jednoduše, uvědomte si, prosím, že nemůžete slova ´přírodní výběr´ používat jen tak svévolně. Předbiologický přírodní výběr je rozpor v pojmech.10
3. Chemická evoluce? To neznáme
Mnoho evolucionistů se bude snažit odmítnout silné důkazy tvrzením, že vznik života z neživých chemikálií nemá nic společného s evolucí a budou tvrdit, že správný termín pro vznik života je abiogeneze. Avšak jejich evoluční kolega Gordy Slack to odmítá, když říká:
- Myslím si, že je lstivé tvrdit, že otázka původu života je pro evoluci nepodstatná. Není o nic méně podstatná, než je velký třesk podstatný pro fyziku či kosmologii. Alespoň teoreticky by evoluce měla vysvětlit celou tu cestu zpět až k tomu úplně nejprvnějšímu organismu, schopnému sebe replikace pomocí biologických nebo chemických procesů. A abychom takovému organismu plně porozuměli, jednoduše potřebujeme vědět, co mu předcházelo. A dnes k tomuto poznání nejsme o nic blíž.11
Zoolog a fyziolog (a evolucionista) Gerald Kerkut (1927-2004) definoval Obecnou teorii evoluce jako „teorii, že všechny živé formy na světě povstaly z jediného zdroje, který sám vznikl z anorganické formy.“12 Samozřejmě, jak bylo uvedeno, nemůže být tato část evoluce nazývána darwinovskou evolucí. Je často zvána chemickou evolucí. Například zářijové vydání Scientific American z roku 1978 bylo zvlášť věnováno evoluci, a jeden větší článek se jmenoval ´Chemická evoluce a původ života´. Říká se tam:
- Zdá se, že to byl J. B. S. Haldane, britský biochemik13, kdo si jako první uvědomil, že pro evoluci života z neživé organické hmoty je nutná redukční atmosféra, tedy taková, kde není volný kyslík.14
Veterán oddaný teoriím o původu života, Cyril Ponnamperuma, je spoluautorem studie se stejným názvem; byl spojen s Laboratoří chemické evoluce, katedry chemie při univerzitě v Marylandu.15 Podobně časopis Discover z roku 2011 publikuje ve svém mimořádném vydání téma „Evoluce: Opětné uvažování nad příběhem života“, kde byl zcela jistě jeden článek o vzniku života.
4. Buňka jako chomáček bílkovin
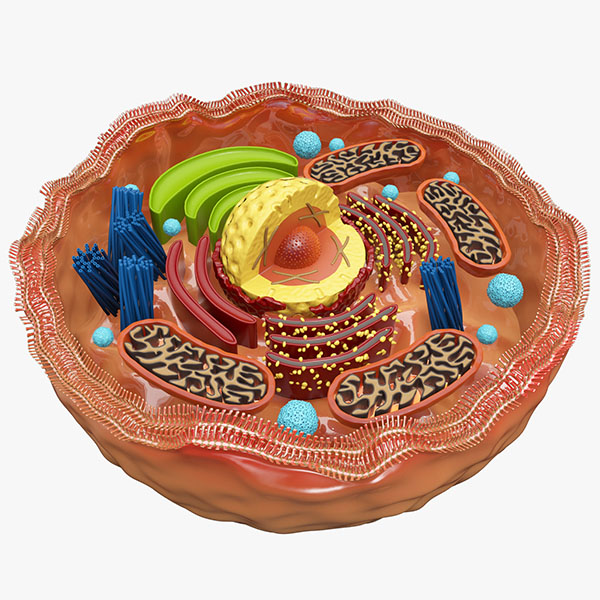
Lidská buňka. Zdroj: www.thing.link.com
Ve dnech Darwina mnoho lidí přijímalo lehkověrně teorii o spontánním plození (samoplození), že život povstal z neživé hmoty. Věřit tomu bylo tehdy o něco snazší, protože struktura buňky byla málo známá. Němec Ernst Haeckel, Darwinův „buldok na kontinentu“ tvrdil, že buňka je jen „jednoduchý chomáček bílkovinné kombinace uhlíku.“16 Pro toto tvrzení však neměl žádnou oporu, protože ještě před dobou Haeckela udělala světelná mikroskopie ohromné pokroky, a to do té míry, že bylo možné rozlišit mnoho jednotlivých buněčných částí.17
Ať tak či onak, molekulární revoluce posledního půlstoletí ukázala, že buňka musí mít jak určitý objem informace, tak i prostředky, jak tuto informaci zpracovat a předat (reprodukovat) další generaci.
4.1. Informace versus chemie
Jak víme, DNA obsahuje ohromné množství kódované informace. Sám ateista Richard Dawkins poukázal na to, že:
- V lidské buňce je dost velká informační kapacita k uložení celé Encyklopedie Britannika, všech jejích 30 svazků, a to až troj či čtyřnásobně.18 Rozdíl mezi životem a neživou hmotou není v látce, ale v informaci. Živé věci obsahují ohromná kvanta informace. Většina této informace je zakódována v DNA…19
Dawkins většinu toho vysvětluje přírodním výběrem, ale jak bylo ukázáno, pro první živou buňku to neplatí. Takže všechna vysvětlování se musejí omezit do oblasti chemie. Jenže v chemizmu samotných stavebních bloků DNA není nic, co by je nutilo spojit se nějakými předem danými způsoby. O nic víc, než síly mezi molekulami inkoustu nutí tyto molekuly spojit se k vytvoření písmen a slov. Michael Polanyi (1891-1976), bývalý prezident fyzikální chemie na univerzitě v Manchesteru (UK), který se začal věnovat filozofii, potvrdil toto:
- Tak, jako uspořádanost potištěné stránky naprosto vzájemně nesouvisí s chemizmem této potištěné stránky, tak také pořadí bází v molekule DNA nemá nic společného s chemickými silami působícími v molekule DNA. Je to právě tato fyzikální neurčitost takovéto sekvence, co vytváří nepravděpodobnost jakékoliv konkrétní sekvence, a tím pádem jí to umožňuje mít význam – význam, který má matematicky vymezený informační obsah.20
Abych to vysvětlil: informace v knize není založena na vlastnostech molekul inkoustu, který je na papíru (nebo pixelech na monitoru, pokud toto čtete v elektronické podobě), ale na způsobu, jakým jsou uspořádány do písmen, slov, výrazů, vět a odstavců. Rozlitá kaluž inkoustu nevytvoří Shakespearovy hry! Všimněte si také, že písmena nedávají žádný smysl, pokud nerozumíte jazyku, ve kterém jsou psána. Například v angličtině slovo ´gift´ znamená ´dar´, ale v němčině toto slovo znamená ´jed´. Špatná domluva může vyústit ve špatné sdělení. Jeden můj německý přítel mi řekl, že když poprvé přijel do země, kde se mluvilo anglicky, pomyslel si, že jsme asi dost pěkní šílenci, když máme všude tolik obchodů s jedem.
Klíčem k životu je informace, ne shoda okolností. Informace není nahodilá (maglajz), ani není zákonitě předvídatelná (spočitatelná), jak je tomu u krystalů, které obsahují jen málo informace. Přední evoluční badatel v oblasti původu života ve 20. století, Leslie Orgel (1927-2007), toto potvrzuje:
- Živé věci jsou rozlišitelné svojí specifickou složitostí. Krystaly, jako u žuly, nesplňují podmínky života, protože postrádají složitost; směsi libovolných polymerů také nesplňují podmínky života, protože postrádají specifitu [vyhraněnost].21
Informace je tedy neperiodické, nepředvídatelné uspořádání signálů, které lze číst a chápat jen systémem, k tomu předem navrženým [zkonstruovaným] a má předem určenou sadu pravidel pro uchovávání, získávání a interpretaci příkazů. Ve všech svých moderních formách je informace vytvářena inteligentním činitelem za účelem zprostředkování sdělení. A život je založen na informaci. Jak říká fyzik a evolucionista Paul Davies (1946- ):
- Nyní víme, že tajemství života nespočívá na chemických složkách jako takových, ale na logické struktuře a organizačním uspořádání molekul…Jako superpočítač, život je soustava na zpracování informací. … Skutečným tajemstvím živé buňky je její software, ne hardware.22
K vyřešení tohoto tajemství však nedělá nic. Místo toho pokračuje: „Jak si hloupé atomy spontánně samy napsaly svůj software?“ Odpovídá: „To nikdo neví …“ …a připouští: „Neexistuje žádný známý fyzikální zákon, který by byl schopen vytvořit informaci z ničeho.“
Pro evoluční teorii je původ informace velkým rébusem. Jednoduše, k jejímu vzniku neexistuje jakákoliv přirozená analogie. Nahodilost je protiklad informace. Stejně jako přesná pravidelnost (jako u krystalů). Aby tedy informace, která není ani nahodilostí ani bezmeznou opakovatelností, mohla vzniknout naturalisticky bez existence mysli či naprogramované soustavy, musely by být překonány tak ohromné pravděpodobnostní bariéry, že to lze oprávněně nazvat ´nemožným´.
Alex Williams toto ještě rozvedl a aplikoval to na buněčný mechanizmus:
- Polanyi poukázal na to, že v živých organizmech jsou konstrukce podobné strojům … A tak, jako konstrukci a funkci těchto běžných částí strojů nelze vysvětlit jen z vlastností kovu, ze kterého jsou utvořeny, tak ani konstrukci a funkci shodných součástí života nelze zredukovat na vlastnosti uhlíku, vodíku, kyslíku, dusíku, fosforu, síry a stopových prvků, ze kterých sestávají. Je nekonečně příkladů těchto neredukovatelných struktur v živých soustavách, ale všechny pracují pod sjednocujícím principem ´autopoiéze´ [„tvorba sebe sama“].23
4.2. A ještě jednou Williams:
- Autopoiéze je jedinečnou a udivující vlastností života – něco podobného neexistuje v celém známém vesmíru. Je tvořena hierarchií neredukovatelně uspořádaných úrovní. Ty zahrnují: (1) součásti s dokonale čistým složením, (2) součásti s vysoce specifickým uspořádáním, (3) součásti, které jsou funkčně propojené, (4) procesy kompletně řízené informacemi a (5) inverzně příčinné metainformační strategie pro přežití jednotlivce i druhu … Každá úroveň je postavena na úrovni pod ní, ale z jejích vlastností nemůže být vysvětlena. A mezi základní úrovní (dokonale čistého složení) a přirozeným prostředím existuje nepřeklenutelná propast.
Takže specifický tvar biomolekul nemůže být předpovídán z vlastností vodíku, kyslíku a uhlíku. Stejně, jako nelze předpovědět pořadí písmen DNA z chemických vlastností těchto jednotlivých písmen. Na každé úrovni existuje nepřeklenutelná hlubina. Williams poukazuje na každou tuto hlubinu jako na Polanyiho nemožnost a podrobně rozpracoval mnoho takových hlubin mezi živými a neživými soustavami. Myslet si, že přirozený svět nějakým způsobem našel kolem těchto hlubin okliku, je podobné víře, že by se všechny vodíkové atomy mohly náhle vyrovnat do přímky od jednoho konce vesmíru ke druhému – jen tak náhodou. To se zcela jistě nikdy nestane, i když je to teoreticky možné. V tomto spočívá základní myšlenka Polanyiho nemožnosti.
5. Hádanka „kuře nebo vejce“ trvá

Kuře, nebo vejce…
Je pozoruhodné, že dekódovací mechanizmus, který „jazyk“ DNA překládá, je sám o sobě zakódovaný v DNA, což je onen známý problém: co bylo dřív, „kuře nebo vejce?“ Vyhlášený filozof vědy Sir Karl Popper (1902-1994) poukázal na to, že:
- To, co činí z původu života a genetického kódu zneklidňující hádanku, je toto: genetický kód nemá žádnou biologickou funkci, dokud není přeložen; čili dokud tento kód nevyústí v syntézu proteinů, jejichž uspořádanost tento kód obsahuje. Ale … mechanizmus, kterým buňka … kód překládá, je sestaven nejméně z padesáti makromolekulárních složek, které jsou samy zakódovány v DNA. Takže kód nemůže být přeložen, nejsou-li použity produkty jeho vlastního překladu. Tohle představuje nepochopitelný kruh; skutečně bludný kruh, jak se zdá, pro jakýkoliv pokus zformovat model nebo teorii vzniku genetického kódu. Takže jsme nejspíš vystaveni možnosti, že se původ života (podobně jako původ fyziky) stává nepřekonatelnou bariérou pro vědu, kdy se všechny pokusy redukovat život na chemii a fyziku stávají nepoužitelným odpadem.24
I když toto Popper napsal před čtyřiceti lety, dnes původ genetického kódu zůstává stejnou záhadou jako tehdy. Nově to připustil i Dawkins v roce 2009:
- „Catch-22“ [Novela Hlava 22 plná paradoxů či bezvýchodná situace] ohledně vzniku života je toto: DNA se může replikovat, ale pro katalýzu tohoto procesu potřebuje enzymy [proteiny]. Proteiny mohou katalyzovat tvorbu DNA, ale k určení správného pořadí aminokyselin potřebují DNA.25
Původ genetického kódu je skutečně bludným kruhem: ke čtení DNA jsou potřeba proteinová zařízení (stroje), ale pokyny ke stavbě těchto proteinových strojů jsou samy o sobě zakódovány v DNA. Navíc používají energii, která potřebuje ATP, jež vzniká pomocí nanomotoru ATP syntáza. A samozřejmě, ten je rovněž zakódován v DNA a dekódován pomocí zařízení vyžadujících ATP! Proteiny jsou zařízení a DNA je materiál pro reprodukci, avšak obojí jsou potřeba v ten samý čas, aby mohla buňka vůbec fungovat. A ovšem, kdyby nebyla žádná informace k reprodukci, bylo by to k ničemu.
6. Dekódování DNA je úchvatný proces
6.1. Transkripce a upínací zařízení
Dokonce i jen prvotní kopírování přesného úseku DNA do mRNA, kódujícího proteiny, vyžaduje velmi sofistikované zařízení. Patří k němu enzym nazývaný RNA polymeráza, který se skládá ze čtyř proteinových řetězců. A je třeba ještě další protein, který určí RNA polymeráze, kde má začít číst templát DNA. Enzymový komplex se pak přesouvá podél řetězce DNA a vždy přiřadí odpovídající RNA písmena, jedno po druhém, než se zastaví přesně na tom správném místě.
Richard Ebright se svým týmem z univerzity Rutgers objevil v tomto procesu transkripce ještě další jemné složitosti.26 Ano, je to právě tato transkribovaná mRNA, která je pak překládána do proteinů ve složitých zařízeních známých jako ribozomy.
DNA jsou dva řetězce a jen jeden je kopírován; pro kopírování se tedy musí rozvinout. Kopírovací zařízení, které se nazývá RNA polymeráza (RNAP), se nejprve naváže na startovní pozici v genu (čili na začátek sekvence kódující protein). Ukotvená RNAP pak vtočí DNA – upnutí.27 Tím se rozvine dvojitý řetězec, aby z jednoho z nich mohla být vytvořena kopie mRNA. Odvíjení také dává energii; je to obdoba navíjení gumy pro pohon vrtule u dětského letadélka. A stejně jako u této hračky, tak i tato energie je pak uvolněna a zařízení se odblokuje a rozjede ze svého startovního místa vpřed. Tím se také znovu natáhne odvinutá DNA (´uvolnění´), která pak uniká ze zadní části tohoto zařízení.
6.2. Translace a ribozom
Ribozom je životně důležité zařízení v buňce, které čte informaci z mRNA a mění na protein. I u tak ´jednoduché´ bakterie, jako je E. coli, žijící ve vašich střevech, se ribozomy skládají z 50 různých proteinů a tří odlišných ribozomálních RNA (rRNA). U složitějších organizmů, které mají v buňce jádro (eukaryotů), je 73 různých proteinů a 4 rRNA. Jeden odborník řekl:
- Ribozom se všemi svými doplňky je asi to nejsofistikovanější zařízení, které bylo kdy vytvořeno. Všechny jeho části jsou aktivní a pohyblivé a je šetrný k okolí; produkuje jen GDP a fosfát.28
Ribozom také zajišťuje, že protein roste lineárně. Mimo toto zařízení by peptidový řetězec snadno vytvářel nežádoucí postranní větve, kde boční skupiny reagují jedna s druhou (třeba aminokyseliny asparagová a glutamová mají větev –COOH, které mohou reagovat s větví –NH2 lysinu nebo argininu). Při průmyslových syntézách peptidů musí být postranní skupiny blokovány inhibičními skupinami, po dokončené syntéze jsou odstraněním těchto skupin odblokovány. Ale u nějaké hypotetické pravěké chemické polévky nebyli žádní organičtí chemici, aby toto prováděli v ty správné okamžiky. Tím je naturalistický vznik života ještě problematičtější, zvážíme-li další aspekty skutečné chemie. Život je postaven na molekulách (DNA, RNA a proteinech), které nejsou vytvářeny někde mimo zcela speciální, uspořádané soustavy (chemické továrny včetně živých buněk).
Bez ribozomů neexistuje žádný život. Není ani žádná myslitelná možnost života bez ribozomů, protože základy chemie blokují opakovatelný vznik proteinů bez soustav, ve kterých probíhá složitý řídící proces, což pro veškerý život představují právě ribozomy.
6.3. Transferová RNA
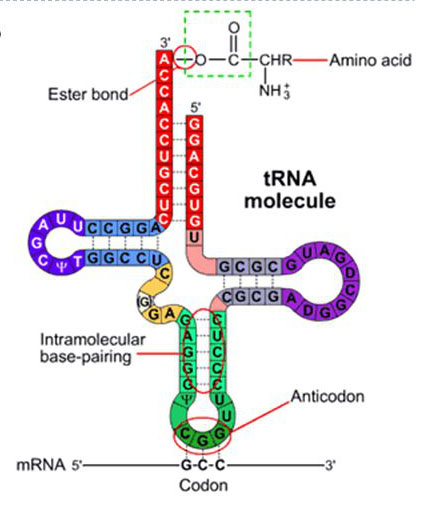
Tranferová RNA. Foto: Wikipedia.com
Molekuly transferové RNA (tRNA) jsou důležité adaptéry, které mají tvar jako čtyřlístky jetelů; mají na starosti nést jednotlivé aminokyseliny do místa, kde jsou přidávány do nového proteinu. Skládají se asi z 80 nukleotidových písmen, z nichž tři se nazývají antikodon. Antikodon má svůj komplementární třípísmenný kodon na mRNA. Tak tRNA přidávají správné aminokyseliny na správné místo v narůstajícím peptidovém řetězci.
Každá aminokyselina musí být také aktivována pro překonání energetické bariéry, která přirozeně brání jejímu napojení na sousední aminokyselinu v roztoku. Pro tento proces dodává energii ATP (o tom bude pojednáno níže). Poté speciální enzym, nazývaný aminoacyl-tRNA syntetáza (aaRS), naváže ve dvou krocích každou aminokyselinu na správnou tRNA. Musí být nejméně 20 aaRS, alespoň jedna pro každý typ aminokyseliny. Jakékoliv selhání by zde zničilo sdělení a tím celý účel genetického kódu.
Molekuly tRNA adaptérů musí mít tu přesně správnou geometrii, aby 1) udržely aminokyseliny v poloze, kde mohou vytvořit peptidovou vazbu a 2) umístily antikodon na správné místo v mRNA. Správná geometrie adaptérů funguje pouze ve spojitosti s ribozomem (buď by se musely vyvinout společně, což má tolik problémů, že to hraničí s nemožností, anebo byly společně zkonstruovány vynikajícím Projektantem). Všechny navrhované scénáře života před existencí ribozomů by nepotřebovaly žádnou specifickou geometrii, takže kde by se tu tato specifická geometrie vzala? Stejně tak je to s hypoteticky se vyvíjejícími adaptéry, které by zřejmě reagovaly každý s každým i s mRNA.29 Je to právě ribozom, co dává buňce kontrolované a řiditelné prostředí se specifickou geometrií, kde lze provádět jemné, avšak nutné úkoly.
Navíc adaptéry tRNA musí být oddělitelné poté, kdy aminokyselina byla připojena na konec rostoucího proteinu. Ribozom se posouvá po mRNA jako po ozubené tyči a energie pro odpojení je dodávána z jiné molekuly pro uchování energie, GTP (guanosin trifosfát), který je postupně produkován složitým, úzce propojeným a regulovaným zařízením.30
Jasnou zprávou je, že toto vše představuje neuvěřitelně složitou soustavu. Jak by něco takového mohlo povstat náhodou, z nějakých nahodilých chemických reakcí v pravěké chemické polévce? Tomuto vyvíjejícímu se systému by se postavilo do cesty velké množství omezení a při porozumění alespoň základům chemie můžeme proti takovému procesu vznést velké množství námitek.
6.4. Dvojitá síta: pokročilá chemická zařízení
Aminoacyl tRNA syntetázy (aaRS) jsou speciální enzymy (proteinová zařízení) vykonávající úžasné chemické reakce. Dokonce od sebe dokážou rozeznat aminokyseliny, které jsou chemicky velmi podobné (jako je např. leucin a isoleucin, které je i v laboratoři velmi obtížné izolovat).31 Jenže i tento malý rozdíl může v důležitých biomolekulách představovat rozdíl mezi životem a smrtí.
Přesto aaRS pro isoleucin je dokáže rozlišit extrémně dobře, s chybovostí jen 1 na 40 000. Dosahuje toho mechanizmem dvojitého síta: jedno síto vyloučí příliš velké aminokyseliny a druhé síto ty příliš malé.32
6.5. Chaperony
Ani proteinový řetězec, který se vytvoří na ribozomu, není ještě hotovým produktem. Aby mohl v buňce vykonávat svoji funkci, včetně enzymů zmiňovaných níže, musí být protein správně sbalen do svého složitého třírozměrného tvaru. Výsledná proteinová konfigurace, která je důsledkem určité sekvence DNA, je určována především buněčnými nástroji zvanými chaperoniny, které jsou samy o sobě proteiny ve tvaru soudku, pomáhající dalším proteinům ve sbalování.33 Bez chaperoninů by se důležitý protein mohl sbalit špatně třeba do smrtícího prionu. Toto je pravděpodobná příčina zhoubného stavu v mozku u Creutzfeldt–Jakobovy nemoci a bovinní spongiformní encefalopatie (BSE), známé pod názvem nemoc šílených krav.
Chaperoniny také omezují evoluční ´pokrok´, protože odmítají špatně sbalené proteiny, a tím omezují počet možných změn. Aby se vyvinul nový protein, musel by nutně procházet tvarovými změnami, z nichž některé působí chaotická řásnění a zprohybání struktury. Musel by však také uniknout nebo se nějak vyhnout chaperoninovým filtrům, které jsou speciálně vyprojektovány, aby se zbavily špatně sbalených proteinů. Tyto nástroje pro konformaci proteinů jsou v živých organizmech všudypřítomné. Jak by kdysi vyšší organizmy vyvinuly chaperoniny tak, aby se daly použít už na začátku existence života? A jak by se první chaperoniny správně sbalily bez předem existujících chaperoninů?34
6.6. Nejmenší motor na světě: ATP syntáza
Další důležitý enzym pro život je ATP syntáza, který vytváří energetické oběživo těla, ATP, adenosintrifosfát (adenosine triphosphate). Energie je podstatnou složkou života a veškerý život používá ATP jako svoje energetické oběživo.35 A skutečně, všechny živé věci, dokonce i bakterie a archea, mají svoje zařízení (motory) ATP syntázu. To dělá pravděpodobně z ATP syntázy nejrozšířenější proteinový komplex na Zemi.
Lidské tělo ve skutečnosti generuje denně asi tolik ATP, kolik samo váží, což je výkon mnoha triliónů těchto zařízení [ATP syntáz]. ATP je velmi rychle spotřebováván, protože dodává energii důležitým biochemickým reakcím, jako jsou syntéza DNA a proteinů, svalové stahy, doprava živin a nervové impulzy. Organizmus bez ATP je jako auto bez paliva, a některé jedy (např. kyanid) působí tak, že zastaví produkci ATP.
Toto zařízení [ATP syntáza] stlačí dvě složky ATP (ADP a fosfát) o dosti vysoké energii pro vytvoření ATP. Poté vychrlí ATP a připraví se na přijmutí nového ADP a fosfátu. Tento motor se otáčí rychlostí asi 10 000 ot/min a každé otočení vyprodukuje tři molekuly ATP.
Ve skutečnosti jde o dva motory v jednom. Horní polovina (nazývaná F1-ATPáza) má tři části, z nich každá je továrnou ATP. Spodní polovina, Fo,37 je přímo napájená elektrickým proudem, který má kladný náboj (proud protonů) namísto námi projektovaných motorů, které pohání záporně nabitý proud (elektronů).38 Od té doby byl však objeven ještě jeden detail, který se týká způsobu, jak jsou tyto motory spojeny a sestaveny.39 Nedávné práce ukazují, že tohle je také ten nejúčinnější motor na světě – je ve skutečnosti tak účinný, jak to jen dovolují zákony fyziky. Výzkumníci dochází k tomuto závěru: „Naše výsledky naznačily 100 % účinnost přenosu volné energie a těsné mechanicko-chemické propojení F1-ATPázy.“40 Je to takový malý div, že i tak přísně světský časopis, jakým je Nature, nazval tyto motory „Opravdové stroje stvoření.“41
7. Bez enzymů není život
ATP syntáza a aminoacyl-tRNA syntetáza jsou jen dva z mnoha typů proteinů, kterým říkáme enzymy. Jsou to biologické katalyzátory, které urychlují životně důležité chemické reakce, aniž by byly samy zničeny. Život vyžaduje enzymy, které jsou extrémně účinné, jinak by nebylo možné přežití, protože bez nich by mnoho podstatných životních procesů probíhalo tak moc pomalu, že by to znemožňovalo existenci života.42
7.1. Superkatalyzátory
Jako dobrý příklad extrémně účinného enzymu ukázal v roce 199843 Richard Wolfenden reakci, která je ´absolutně klíčová´ pro vznik stavebních bloků DNA a RNA, a která by ve vodě trvala 78 miliónů let“,44 zatímco pomocí enzymu46 byla urychlena 1018krát.45 Tento enzym vyžaduje pro svoji funkci speciální, jednoznačně definovanou strukturu.47
V roce 2003 našel Wolfenden fosfatázu, která katalyzuje hydrolýzu (štěpení) fosfátových vazeb, a která zvyšuje rychlost reakce 1021krát, dokonce tisíckrát víc než předchozí enzym! Tento enzym umožňuje, aby se během setiny sekundy odehrály životně důležité reakce, ke kterým patří buněčná signalizace a ovládání. Bez tohoto enzymu by tyto podstatné reakce trvaly bilióny let, téměř stokrát déle, než je údajný evoluční věk vesmíru (kolem 15 miliard let)!48 A ano, tyto enzymy, spolu s dalšími výše vyjmenovanými procesy, musely existovat už v té úplně první replikující se buňce, aby mohla tato buňka přežít a předat svoji DNA, a ta DNA musela kódovat právě ty proteiny, které jsou k těmto procesům potřebné!
7.2. Jaké to má důsledky?
Wolfenden řekl:
- Bez katalyzátorů by od mikroba po člověka neexistoval vůbec žádný život. Dohání vás to k úžasu, jak přírodní výběr fungoval, že vyprodukoval protein, který se odlepil od země jako primitivní katalyzátor pro tak mimořádně pomalou reakci.49
Tohle je však kuriózní slepá skvrna. Jak už bylo dříve řečeno, přírodní výběr nemohl fungovat, dokud zde nebyl život, a na druhé straně, jak on říká, život nemohl fungovat bez těch enzymů, které enormně urychlují životně důležité reakce. Původ života byl od samotného začátku základními zákony chemie odkázán k nezdaru!
8. Nejjednodušší bakterie: 482 genů a 580 tisíc bází
Aby toho nebylo málo, tak spíše než chemie je tím největším problémem vzniku života informace. I ten představitelně nejjednodušší život by musel mít obrovský informační obsah. Nejmenší známý genom ze všech známých organizmů má bakterie Mycoplasma genitalium (vir nepřipadá v úvahu, protože je zcela závislý pro svoji reprodukci a sestavení na aparátu mnohem složitějších buněk).50 Mycoplasma má 482 genů s 580 000 bázemi.51 Samozřejmě, že tyto geny fungují jen za přítomnosti už předem existujícího aparátu pro translaci a replikaci spolu s buněčnou membránou atd. Ale Mycoplasma genitalium nemá žádné buněčné stěny, a může tak přežívat jen parazitováním na složitějších organizmech (např. žije v buňkách respiračního systému a urogenitálního traktu lidí), které jí poskytují mnohé živiny, jež si sama nedokáže opatřit. A opravdu, tento organismus vznikl patrně ztrátou genetické informace, což ho učinilo závislým na hostiteli.52
Mykoplazmy jsou velmi jednoduché bakterie (i když pořád ještě velmi složité). Mohlo by vzniknout něco ´jednoduššího´? Badatel Eugene Koonin, v jehož zájmu je vytváření umělých biologických organizmů, se pokusil před deseti lety spočítat to úplně nejnutnější minimum požadavků pro živou buňku. Tuto práci založil na mykoplazmách a odhadl, bez kolika genů se i tyto jednoduché buňky obejdou. Jeho tým dospěl k výsledku 256 genů [jako potřebné minimum pro tuto bakterii].53
Byli na pochybách, zda by tento hypotetický tvoreček mohl dlouho přežít, protože takový organismus si sotva opraví poškozenou DNA, nemůže si dlouhodobě koordinovat schopnosti svých zbývajících genů, bude postrádat schopnost trávit složité sloučeniny a bude vyžadovat komplexní zásobu organických živin ve svém okolí.
Nepřekvapuje, že další výzkum toto číslo významně upravil směrem nahoru. Tento nový hypotetický minimální genom sestává z 387 genů kódujících proteiny a 43 genů kódujících RNA.54
Článek v New Scientist z roku 2009 říká:
- Není pochyb o tom, že společný předek měl DNA, RNA a proteiny, univerzální genetický kód, ribozomy (továrny na stavbu proteinů), ATP a protonově poháněný enzym pro tvorbu ATP. Musel mít též propracovaný mechanizmus pro čtení DNA a převod genů na proteiny. Krátce řečeno, ten poslední společný předek veškerého života vypadá velice přesně jako moderní buňka.55
Když v roce 2011 evoluční biologové diskutovali o tomto hypotetickém Posledním univerzálním společném předkovi (LUCA – Last Universal Common Ancestor), tak si také uvědomili, že to nemohlo být nic jednoduchého. Spíše by měl mít univerzální předek ještě navíc ´univerzální organelu´ k ukládání vysoce energetických sloučenin zvaných pyrofosfáty; podle předchozího dogmatu bakterie organely neměly.56 Zpráva říká:
- Podle zprávy badatelů nová fakta ukazují, že LUCA byl přece jenom sofistikovaný organismus se složitým uspořádáním, jaké pozorujeme u buňky.57
Žádné modelování vzniku života se ani v nejmenším tomuto nutnému minimu nepřiblíží.
8.1. Vznikla tato složitost náhodou? Ani náhodou
Přírodní výběr nemůže fungovat bez samostatného reprodukčního systému. Takže pro vysvětlení i této minimální úrovně složitosti se na něj nelze odvolávat. Vše, co evolucionisté mají k dispozici, je náhoda. A náhoda podléhá docela jednoduchým pravděpodobnostním výpočtům.
Informační teoretik Hubert Yockey propočítal, že kdybychom měli nádrž čistých, aktivovaných biologických aminokyselin (což je o mnoho štědřejší než by obsahovala údajná chemická ´prapolévka´), celkové množství informace, která by mohla být vyprodukována během jedné miliardy let pokusů a omylů, jak to evolucionisté předpokládají, by byl pouze malý jednoduchý polypeptid o délce 49 aminokyselinových zbytků.58 Což je asi kolem 1/8 velikosti (a tím pádem i obsahu informace) běžného proteinu, avšak ona hypotetická jednoduchá buňka, jak byla výše popsána, potřebuje nejméně 387 proteinů (které jsou všechny předem zakódovány v DNA!), a to by jí pouze umožňovalo žít ve velmi specifickém a neměnném prostředí se stálou zásobou energeticky bohatých živin a biomolekul. A tento Yockeyho odhad velkoryse předpokládá, že mnoho chemických překážek může být překonáno, což je obrovský předpoklad, jak ještě bude ukázáno.
Podobně lze propočítat pravděpodobnost nahodilého získání sekvence DNA pro každý z těchto proteinů. Jistě existuje pro mnoho sekvencí určitá volnost, ne však kolem aktivních míst. I evoluční autoři mlčky připouštějí, že některé sekvence jsou klíčové. Nazývají je ´konzervované´ (tím se míní, že daná sekvence byla tak důležitá, že ji přírodní výběr zakonzervoval vyloučením různých dalších variant, které se v evoluční historii objevily). Konzervované celé proteiny zahrnují histony, které se chovají jako cívky, kolem kterých se obaluje v chromozomech DNA, ubikvitin, který je kromě bakterií v organizmech všudypřítomný a podstatný pro označování proteinů určených k likvidaci,59 a kalmodulin, všudypřítomný protein, který váže vápník a jehož téměř všech 140-150 aminokyselin je ´konzervovaných´.
Následující výpočet bude k evolucionistům značně štědrý. Budeme se tvářit, že na jeden enzym je jen 10 konzervovaných aminokyselin a že existuje nějaký mechanizmus ke spojování aminokyselin do dlouhých řetězců v roztoku (to je ovšem až moc štědré, protože voda v roztoku neustále hydrolyzuje peptidové vazby):
- 20 aminokyselin
- 387 proteinů pro ten myslitelně nejjednodušší život
- 10 konzervovaných aminokyselin průměrně
∴ šance je 20-3 870=10-3 870.log20=10-5 035
To znamená jedna možnost ku jedničce následované 5 000 nulami. Takže to je těžší než uhádnout správně PIN, který má 5 000 číslic, na první pokus!60 Jenže bez takového šťastného sebe poskládání je život vyloučený. Zde se nejedná o pomalou postupnou konstrukci chemikálií před ožitím, neboť jde o nejjednodušší formu života podle těch, kteří věří, že život může vzniknout z pravěké chemické polévky. Musíme mít stále na paměti, že buňka musela vzniknout naráz, v jednom okamžiku, jinak by nepřežila nehostinné prostředí a vůbec by neožila. Žádné milióny let trvající vývoj chemikálií není z přírodovědeckého hlediska možný!
Mnoho evolucionistů vyřklo myšlenku, že dáme-li věcem dostatek času, všechno je možné. Je opravdu čas tím ´hlavním hrdinou dne´? Není. Máme:
- 1080 atomů ve vesmíru
- 1012 interakcí atomů za sekundu
- 1018 sekund od vzniku vesmíru podle pochybné teorie velkého třesku
∴ jen 10110 interakcí je k dispozici
Toto je obrovské číslo, ale když ho porovnáme s tím, kolik pokusů by bylo třeba, abychom dostali přijatelnou šanci získat správnou sekvenci nukleotidů, která je potřebná pro kódování proteinů nezbytných pro tu myslitelně nejjednodušší formu života, je toto číslo absurdně nízké. I při těchto reakcích by byla jen jedna šance v 104 925. Tato čísla jsou tak ohromná, že jsou zcela nesmyslná. Jejich smysl je jen v ilustraci statistické nemožnosti (za hranicemi ´nepravděpodobnosti´) vzniku života z neživých chemikálií.
Známý kosmolog Sir Fred Hoyle (1915-2001) opustil svůj ateismus poté, co zvážil tyto absurdně nízké pravděpodobnosti:
- Možnost, že se život zformoval z bezduché hmoty, je jedna k číslici, za kterou je 40 tisíc nul…což je dostatečně velké na pohřbení Darwina a celé evoluční teorie. Neexistovala žádná pravěká chemická polévka, ani na této, ani na jiné planetě, a jestliže tedy počátky života nebyly nahodilé, pak musely být produktem smysluplné inteligence.61
9. Pověsti o molekulách, které se samy replikují
Ve snaze vyrovnat se s pichlavým Popperovým dilematem (viz výše) a požadavkem na minimální složitost života někteří evolucionisté teoretizovali, že jeden typ molekuly mohl mít jak katalytickou, tak i reprodukční roli. Volba většinou padá mezi nukleové kyseliny (RNA) a proteiny.
Jenže nyní musí i evolucionisté připustit, že RNA je ve skutečnosti velmi špatný katalyzátor a proteiny jsou špatné replikátory. Žádný RNA enzym se nepřiblížil podobné účinnosti proteinových enzymů (pro živé tvory nutných), analyzované Dr. Wolfendenem – život jako takový je bez těchto schopností nemyslitelný. Je také zcela bez užitku teoretizovat o nějakých jiných formách života, protože máme vysvětlit ten život, který vidíme na planetě Zemi. A tento typ života popírá naturalistické příběhy o svém vzniku. Evolucionisté připouštějí, že ribozomy (RNA enzymy vytvořené při laboratorních experimentech) nejsou účinné enzymy; nikdy by nemohly dosáhnout účinnosti enzymů potřebných pro život. Podobně i Dawkins musel uznat, že:
- Darwin v odstavci o ´malých teplých jezírcích´ spekuloval, že klíčovým momentem při vzniku života mohl být spontánní vznik proteinu, ale toto se jeví méně slibné než většina Darwinových myšlenek. … Proteiny jsou v jedné věci velmi moc špatné, a to Darwin přehlédl. Jsou zcela beznadějně neschopnými replikátory. Nedokážou kopírovat samy sebe. To pak znamená, že klíčovým krokem ke vzniku života nemohl být spontánní vznik proteinu.63
Existují ještě další ohromné chemické potíže u modelů s první RNA či prvním proteinem. A zastánci jednoho modelu jsou ve skutečnosti nejostřejšími kritiky modelu druhého, takže každá ta strana má ve své kritice pravdu – tudíž, život se zcela jistě nevyvinul z chemikálií!
9.1. Svět ´RNA´
Velmi populární myšlenka, že život byl založen při svém vzniku na RNA, sahá až k roku 1967. Tehdy Carl Woese tvrdil, že RNA nevykazovala jen schopnosti reprodukce, ale mohla též působit jako katalyzátor, a tak plnit obě role.64 Thomas Cech a Sidney Altman nezávisle na sobě předvedli, že některé sekvence RNA mají katalytické účinky. Za „objev katalytických vlastností RNA“ dostali v roce 1989 Nobelovu cenu za chemii.65
Objev takovýchto ribozymů vedl mnohé evolucionisty k vyhlášení Světa RNA. Navrhovali, že první život sestával hlavně z RNA, která se dokázala nejen reprodukovat, ale byla schopna i mnoha funkcí prováděných dnes enzymy. Občas je zastáncem této myšlenky Richard Dawkins.66
Jenže s hypotézou světa RNA je spojeno mnoho problémů:67
- RNA je ve skutečnosti velmi složitá molekula. Tvrzení, že mohla vzniknout v pravěké chemické polévce, je úletem do fantazie.
- RNA je dokonce méně stabilní než DNA, a to DNA je už sama o sobě extrémně nestabilní (viz níže)
- Ale i stavební kameny RNA (nukleotidy) jsou samy o sobě docela složité molekuly, a nemohly vzniknout v pravěké chemické polévce. Vytvořit je v laboratoři vyžaduje docela složitou chemii.
Aby se vytvořil protein, musí se vzájemně spojit aminokyseliny peptidovými vazbami, čímž se vyloučí molekula vody. Je tu však mnohem větší sklon k reverzibilní reakci. O to větším problémem je to ve vodě.
- Pokusy s jiskrovým výbojem, jako byly ty Miller-Ureyovy, nevytvoří RNA/DNA bázi cytosin. Cytosin sám o sobě, i kdyby mohl vzniknout, je příliš nestabilní k tomu, aby se jej nahromadilo dostatečně využitelné množství i v oněch údajných geologických ´dlouhých obdobích´, neboť jeho poločas rozpadu při teplotě 25 °C je 340 let.68
- I ty jednodušší stavební kameny RNA jsou mimo buňku nestabilní. Poločas rozpadu ribózy je pouhých 44 let při pH 7,0 a 0 °C. Při vysokých teplotách je to ještě horší (např. 73 minut při 100 °C).69 A báze RNA jsou velmi rychle zničeny ve vodě horké 100 °C, což je problémem pro ´teplá jezírka´nebo hydrotermální teorie.70 Adenin a guanin mají poločas rozpadu okolo jednoho roku, uracil 12 let a cytosin jen 19 dní.71
- Jak bylo uvedeno výše u aminokyselin, nukleotidy nepolymerizují spontánně; musí být aktivovány. Navíc optimální podmínky pro podporu polymerizace také podpoří odbourání ribozymů a templátů, které jsou potřebné, aby tento proces fungoval.72 Ještě horší je, že následkem všudypřítomného procesu hydrolýzy (která rozkládá dlouhé řetězce biomolekul) je voda všeobecným nepřítelem polymerizace.73
Dva enantiomery zobecněné aminokysliny, kde R je jakákoliv funkční skupina (vyjma H)
- Chemické reakce, při kterých se tvoří nukleotidy, vytvářejí chirální molekuly (zrcadlový obraz). Jakákoliv uhlík obsahující molekula se čtyřmi postranními řetězci může mít ´levotočivou´ nebo ´pravotočivou´ formu (viz diagram), a jakákoliv reakce v pravěké chemické polévce by vygenerovala směs 50:50 od všech.74 K vytvoření řetězce nukleotidů, který se může sbalit do šroubovice (což je nutné pro stabilitu a replikaci), musí být celý komplex výlučně ze ´stejně točivých forem´ (neboli homochirální). Všech pět nukleotidových bází (A, C, G, T a U) je pravotočivých, stejně jako všechny cukry v páteři DNA. Je zajímavé, že všechny aminokyseliny, které život používá, jsou levotočivé. Následkem požadavku na homochiralitu i jen malá část nesprávně orientovaných molekul ukončí replikaci RNA, a to jak v živých, tak dokonce i v umělých soustavách.75
- I kdyby se takové polymery mohly tvořit, což by muselo nejprve probíhat bez předem existujícího templátu, musely by být pak schopny se replikovat. Tato replikace musí být přesná, jinak by se ztratily jakékoliv informace, které byly získány náhodou. I byť jen 96,7 % přesnost u jednoho hodně uváděného případu,76 je zcela nedostatečná přesnost – výsledkem bude chybová katastrofa. Replikace lidské DNA má míru chybovosti přibližně jedné chyby na každou miliardu replikací, díky velmi dobře vyprojektovanému, sofistikovanému korekčnímu mechanizmu.
- Pokud by sebe replikující molekuly RNA byly základem první živé formy, musely by mít všechny potřebné funkce k udržení (zachování) organizmu. Jenže u RNA nebyla prokázána žádná taková schopnost kromě určitého omezeného počtu chemických reakcí.
- A na vrcholu toho všeho, jak by mohl takový RNA organizmus umožnit vznik modernímu organizmu, s proteinovými katalyzátory zakódovanými v reprodukující se DNA? Tohle vyžaduje zcela úplně novou úroveň dekódovacího zařízení77 a vyžadovalo by to zcela nepravděpodobné přepnutí od RNA k DNA/informaci pro ukládání a použití založeném na proteinech.
- Protože je fosfát základní složkou nukleových kyselin, musí zde být neodmyslitelný volný fosfátový iont (PO43–), jenže badatelé ´světa RNA´ používají typickou, asi milión násobnou koncentraci ze současného moře. V přírodě se fosfát rychle vysráží v důsledku přemíry vápníkových (Ca2+) a hořčíkových (Mg2+) iontů, takže dojde k jeho pevné vazbě v nerozpustných pevných látkách, a to při všech známých i teoretických scénářích.78
Není divu, že jeden z předních výzkumníků v oblasti modelování ´světa RNA´, Gerald Joyce, napsal:
- Nejrozumnějším předpokladem je, že život nezačal s RNA. … Přechod na svět RNA, stejně jako vznik života obecně, je zatížen nejistotou a sužován nedostatkem experimentálních údajů.79
Poté, co jiný evoluční chemik, Robert Shapiro ukázal, že jeden ze stavebních kamenů RNA je nehodnověrnou složkou pravěké chemické polévky, uvedl:
- důkazy, které jsou v současné době k dispozici, nepodporují myšlenku, že by RNA nebo jiný alternativní replikátor, používající sadu současných bází RNA, byl přítomen v začátcích života.68
Nedostatek jakýchkoliv funkčních scénářů pro původ života je bolestnou stránkou evoluční teorie. Není to tak, že bychom něco opomněli. Není to tak, že bychom snad pořád ještě něčemu nerozuměli. Zatímco ty věci jsou jistě správně, všechno, co jsme se až dosud naučili o fyzice, chemii a pravděpodobnosti, ukazuje jinam než k původu života z anorganických chemikálií.
10. Bajky o prvotních proteinech
Starší teorie chemické evoluce navrhovala, že tu nejprve byly proteiny. Tato myšlenka zažila intenzívní mediální propagandu po Miller-Ureyových pokusech v padesátých letech 20. století.
10.1. Miller-Ureyovy pokusy
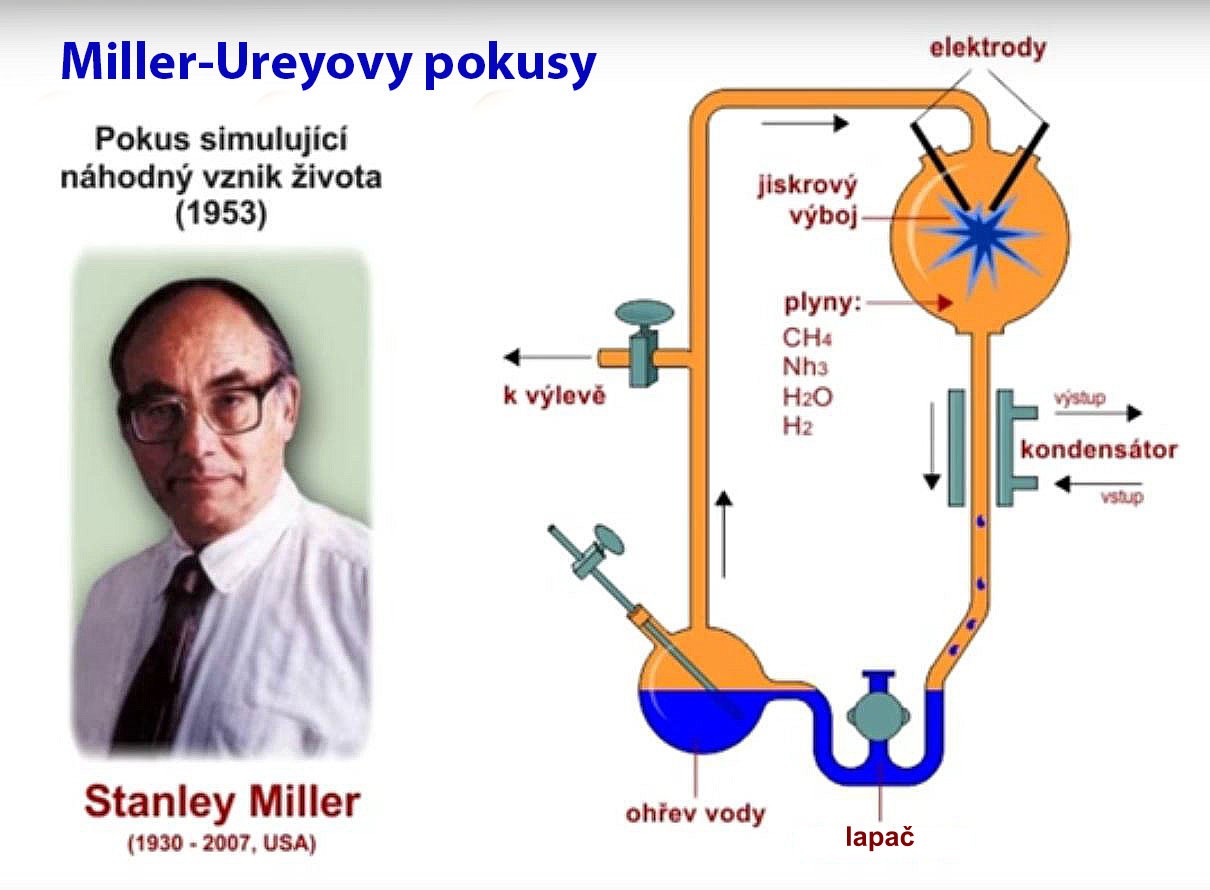
Miller-Ureyův experiment
Stanley Miller (1930-2007) byl postgraduálním studentem Harolda Ureye (1893-1981), který získal v roce 1934 Nobelovu cenu za chemii za objev deuteria (těžký vodík).80 Miller a Urey naplnili uzavřenou skleněnou aparaturu redukčními plyny metanem, amoniakem a vodíkem (ale vyloučili oxidující plyny jako je kyslík). K tomu přidali nádobu s vřící vodou pro přísun vody a k zajištění cirkulace plynů proudících přes jiskrový výboj o 60 000 voltech, což mělo simulovat atmosférické výboje (blesky). Směs plynů také procházela vodou chlazeným kondenzátorem, aby tak jakékoliv produkty reakcí byly shromážděny tím, že klesly do níže umístěné vodní pasti.
Po týdnu zjistili ve studené pasti červenou skvrnu. Skládala se z velké části z nerozpustné, jedovaté a karcinogenní směsi nazývané dehet nebo pryskyřice, což je běžný produkt organických reakcí. V tomto dehtu zjistili malé množství aminokyselin.81
To však byly hlavně nejjednodušší aminokyseliny glycin a alanin.82 A výtěžkem bylo jen nepatrných 1,05 % a 0,75 % daných produktů. Miller připustil, že „Celkový výtěžek je nízký vzhledem k vynaložené energii.“82 Není však žádný důkaz, že by se situace v delším časovém úseku zlepšila. Tyto nepatrné výtěžky jednoduchých produktů se zdají být maximem toho, čeho lze vůbec kdy u této simulace dosáhnout.83
Jaká ironie probírat se všemi těmi domněnkami, výsledky opakovaných pokusů nebo i důmyslně postavených zařízení, aby bylo docíleno nepatrného výtěžku několika málo aminokyselin, které v žádném ohledu nemohou vytvořit život, a přitom všem prohlašovat, že ´pro vznik života nebyla potřebná žádná inteligence´! Miller a Urey ve skutečnosti přišli na to, že nahodilé chemické reakce produkují nahodilé chemikálie. A tyto chemické produkty byly stejně tak jednoduché. Nahodilá sbírka jednoduchých molekul je pravým opakem toho, co je pro život nezbytné.
11. Katem samovolného vzniku života je sama chemie
Informační molekula v buňce, DNA, je opravdu velmi nestabilní molekula. Nedávná studie o stabilitě DNA odhaduje, že i při uložení v kosti, by se DNA úplně rozpadla za 22 000 let při 25 °C, za 131 000 let při 15 °C a za 882 000 let při 5 °C; a za 6,83 miliónů let by se DNA rozpadla při -5 °C.84 Jiný článek uvádí:
- „Existuje všeobecně rozšířená víra, že DNA je ´pevná jak skála´- extrémně stabilní,“ říká Brandt Eichman, mimořádný profesor biologických věd ve Vanderbiltu, který tento výzkum řídil. „Ve skutečnosti je DNA vysoce reaktivní.“85
Kolem jednoho miliónu bází je denně zničeno v DNA každé lidské buňky. Tato poškození jsou způsobena kombinací více činitelů: běžnou chemickou aktivitou uvnitř buňky, vlivem záření a jedovatých látek z prostředí, včetně cigaretového kouře, grilovaných jídel a průmyslových odpadů.86
Kvůli této vysoké úrovni každodenního poškození, musí mít živé bytosti důkladný mechanizmus pro opravy DNA. Biolog James Shapiro z Chicagské univerzity poukázal na to, že:
- Všechny buňky od bakterie až po člověka mají opravdu úžasnou paletu reparačních systémů, které slouží k odstraňování druhotných a stochastických zdrojů mutací. Víceúrovňové kontrolní mechanizmy rozpoznají a odstraní chyby, které se během replikace DNA nevyhnutelně vyskytnou … buňky se chrání právě přesně těm druhům náhodných genetických změn, které podle klasické teorie jsou zdrojem evoluční variability. Tím, že živé buňky mají schopnost kontrolovat a opravovat svoje vlastní systémy, nejsou pasivními oběťmi nahodilých sil chemie a fyziky. Buňky věnují velké zdroje potlačování nahodilých genetických variací a mají schopnost určit si na svém pozadí úroveň mutability tím, že si seřídí aktivitu svých vlastních reparačních systémů.86
Samozřejmě, že hypotetická pravěká chemická polévka neobsahovala tyto úžasné opravné systémy. Tím pádem, i kdyby se byla DNA schopna sama od sebe nějak zformovat, dlouho by nepřežila.
RNA je ještě víc nestabilní než DNA. Tato nestabilita RNA je hlavním důvodem, proč mnoho zastánců chemické evoluce dává přednost scénářům, kde všechno začalo proteiny. Jenže rovněž i proteiny jsou v delším časovém měřítku velmi nestabilní, a tak by byly stejně tak zničeny. Jsou přece podrobeny druhému zákonu termodynamiky, takže se nakonec rozpadnou následkem nahodilého pohybu atomů a záření z prostředí. Například voda má tendenci proteiny rozložit na jejich základní složky, což jsou aminokyseliny (hydrolýzou),87 a budou podrobeny destruktivním křížovým reakcím s dalšími chemikáliemi88 v předpokládané pravěké chemické polévce.89 A podobně jako nukleotidy, o kterých byla řeč výše, budou i aminokyseliny vytvářeny jako směs pravo a levo točivých forem (racemáty), spíše než to, co je pro biologické enzymy skutečně potřebné, a to je homochiralita (pouze čisté pravotočivé nebo čisté levotočivé formy).90
Poslední odhady udávají pro kolagen horní limit 2,7 Ma (miliónů let) a pro kostní protein osteokalcin 110 Ma při bodu mrazu (0 °C). Při mírnějším chladu 10 °C jsou horní limity mnohem nižší – 180 000 let pro kolagen a 7,5 Ma pro osteokalcin. Při teplotě 20 °C jsou maximální životnosti ještě kratší: 15 000 let a 580 000 let.91 Protože se zpravidla rychlost reakcí exponenciálně zvyšuje s teplotou (biologické reakce se většinou zdvojnásobují každých 10 stupňů Celsia),92 stává se tento problém pro teorii teplých jezírek neřešitelný. Nestabilita proteinů je skutečně velmi silným argumentem proti dlouhým věkům: nalézáme je v dinosauřích kostech, ale pokud by tyto kosti byly opravdu >65 miliónů let staré, už by v nich nezůstal žádný protein.93 Tento problém se pro evolucionisty ještě zhoršil, když byla v dinosauří kosti nalezena DNA. Pokusy vyloučily kontaminaci a DNA byla natolik nepoškozena, že tvořila malé neporušené dvoušroubovice.94
12. Psst! Bez plánu ani ránu
Modelování vzniku života chce ukázat, že život mohl vzniknout v důsledku času a náhody. Ve skutečnosti i ty chabé výsledky mohly být dosaženy jen proto, že celý pokus byl naplánován. K běžnému postupu patří, že se hledá stopa sloučeniny A v pokusu s jiskrovým výbojem, a sloučeniny B v jiném modelu (někdy při vzájemně neslučitelných podmínkách), a pak se řekne „Podívejte se, A a B vzniknou za realistických podmínek na prvotní zemi.“ Poté se od průmyslové chemické společnosti získá homochirální koncentrované A a B a nechají se zreagovat, aby vytvořily stopové množství složitější sloučeniny C.95 Načež zprávy vytrubují, že se C vytvoří za podmínek na prvotní zemi. Toto však neukazuje, že zředěné A a B může spolu reagovat a zároveň také i společně vzniknout, a že nebudou reagovat s nečistotami D, E nebo F, které se také vytvořily při prvních pokusech. Krátce řečeno, evolucionistické modelování je zatíženo nepřípustnou úrovní promyšlených zásahů.96
Mnoho z evoluční propagandy se podobá následující hypotetické teorii o vzniku automobilu:
- Protože je projekt (design) nevědecké vysvětlení, musíme tedy místo toho najít nějaké naturalistické (přírodní) vysvětlení. Pokusy bylo prokázáno, že jedna důležitá stavební jednotka auta – železo – může vzniknout zahříváním přírodních minerálů, jako je hematit, na teploty, které se na některých místech Země vyskytují. A co víc, bylo prokázáno, že železo může pod tlaky vytvářet tenké pláty, a tyto tlaky se v některých geologických formacích vyskytují. …
Pokud by se snad tento spontánní vznik auta jevil někomu jako za vlasy přitažený, poznamenejme, že i ta nejjednodušší možná buňka má o mnoho vyšší informační obsah než automobil, který nemusí udržovat nějakou vnitřní homeostázi, natož se sám reprodukovat.
12.1. Abiotická chemie – protiklad života
Naše biomolekuly nezískávají svoje vlastnosti ze svých komponentů, per se, ale řečeno přesně, z uspořádanosti těchto komponentů, tedy z informace. Tato uspořádání nevznikají z fyzikálních a chemických vlastností těchto komponentů, ale jsou do nich vložena z vyšší úrovně. Zopakuji-li to, co už bylo řečeno, vlastnosti uhlíku, vodíku, kyslíku, dusíku, fosforu a síry nelze použít k predikci určitého makromolekulárního tvaru (je zde nekonečný počet možností). Je to spíše tak, že makromolekuly byly navrženy vykonávat určité funkce s ohledem na vlastnosti těchto jednodušších prvků. Makromolekuly byly vyprojektovány shora dolů; nevyvinuly se ze svých základů na svůj vrchol. V živých bytostech již předem přítomná uspořádanost daného mechanizmu (informace) ukládá tuto uspořádanost na jednotlivé komponenty, aby vytvářely další kopie, přičemž toto zařízení vzniklo stejným způsobem z předchozích generací, a tak to pokračuje dál a dál. Mechanizmus nepovstane ze svých komponentů stejně tak, jako izoenergetický šum (bílý šum) nevytvoří symfonii.
Čím více rozumíme biochemii, tím více poznáváme, jak je odlišná od abiotické chemie. Zákony jsou ty samé, ale chemismus vně živé buňky je vždy ´špinavým´ chemickým působením hmoty, zatímco biochemie je chemií jednotlivých čistých molekul.97
Termín ´špinavá´ pochází od evolučního chemika a laureáta Nobelovy ceny, Christiana de Duve. Je to pouhé konstatování faktu, že chemismus neživého světa je tvořen současně mnoha molekulami a pravidelně obsahuje kontaminanty.98 Teistický evoluční paleontolog Simon Conway Morris nazýval produkty typických pokusů se vznikem života „blátem“, „maglajzem“ a „svinstvem“,99 v souznění s evolučním chemikem Grahamem Cairns-Smithem, který používal výraz „hrubě znečištěný sajrajt“.100
Zůstává pravdou, že moderní průmyslová chemie se svými pečlivě naplánovanými procesy nemůže běžně dosáhnout čistot nad 99,99 %. Je to ponejvíce proto, že pracuje s obrovským počtem molekul najednou. Ale biochemie může dosáhnout lepších výsledků, protože každý enzym pracuje ve stejné době jen s jednou molekulou. Další enzym, ve správném tvaru a na správném místě, se pak o tyto produkty postará.
Závěrem, chemismus buňky je přesný, v pevných mezích, řízený a plně funkční, a bez toho nemůže existovat ten život, který známe. Chemie mimo buňku je opakem toho, co je třeba k vytvoření života. Abiotická chemie je protikladem života. Jak potom mohla vyprodukovat první živou buňku?
12.2. Výzkumníci balili pokusy do měkké peřinky
Všechny zdroje energií, které produkují biochemické látky, je ještě mnohem rychleji ničí. Už jsem zmínil hydrolyzující účinek vody, který má snahu mnohem rychleji rozrušit peptidové vazby, než jak se v roztoku tvoří. Energetické zdroje působí také mnohem destruktivněji než konstruktivně. Například na hypotetické pradávné zemi by existovaly dvě formy UV záření. Jedna z nich je destruktivní (ionizační), krátkovlnné ultrafialové (UV) světlo. Druhé je nedestruktivní UV o delší vlně. Dlouhovlnné UV může být v některých případech konstruktivní. Např. může být pohlcováno a využíváno chlorofylem.101 Jenže ona destruktivní forma UV záření je hojnější než ta konstruktivní, a je také mnohem účinnější. To má dvojnásobný dopad, takže destruktivní účinky jsou oproti konstruktivním asi 104-105krát silnější.102
Miller-Ureyovy pokusy používaly strategicky navržené pasti, aby byly biochemické látky izolovány hned, jak se vytvořily, aby je nezničily jiskry (nebo v pozdějších experimentech používané UV záření). Bez těchto pastí by nepřežilo ani to nejmenší množství, které bylo získáno. To však není realistické napodobení pravěké chemické polévky, ve které nebyly žádné přijatelné prebiotické mechanizmy pro záchranu aminokyselin před UV zářením, které by tyto produkty rychle stáhly z atmosféry, a také před vystavením účinků vody. Mějme na mysli, že člověk může být v některých případech ošklivě spálen od slunce i za podmračeného dne, a dokonce i pod vodou, neboť UV záření proniká až desítky metrů čistou tekutou vodou.103 Takže tyto pasti byly jen dalším příkladem nepřijatelné úrovně zasahování inteligentními výzkumníky.
13. Máme scénář chemické evoluce. Tak co tu chcete dokazovat?
Informační teoretik Hubert Yockey, který není kreacionistou, publikoval před 30 lety tento velmi zasvěcený komentář:
- „Výzkum vzniku života je unikátní v tom, že jeho výsledek už byl autoritativně přijat … . Co ještě zbývá udělat je objevit takové scénáře, které by detailně popsaly mechanizmy a procesy, za kterých ke vzniku života došlo.“104
Tohle je dobré mít na mysli při četbě populárních popisů evoluce, nebo když reagujeme na tvrzení, že lidé, kteří věří, že život byl vyprojektován, jsou ´zaslepení´. Dr. Yockey svoji studii zakončil slovy:
- Lze to uzavřít tak, že v protikladu k pevně založenému a normálnímu zdravému rozumu, ještě nebyl popsán scénář, který by vysvětloval genesi života na Zemi pomocí náhody a přírodních příčin, a který by mohl být přijat na základě faktů, nikoli víry.105
Nadace pro Vznik života nabízí v současné době cenu jednoho miliónu dolarů komukoliv, kdo přijde s chemicky přijatelným řešením samovolného vzniku genetického kódu a života. Jejich webová stránka uvádí:
- ´Odměna Původ života´ ® (nadále nazývaná „Odměna“) bude vyplacena za nabídku vysoce přijatelného mechanizmu pro spontánní vznik genetických instrukcí v přírodě, které by byly dostačující ke vzniku života. Pro získání této ceny musí být vysvětlení shodné s empirickými biochemickými, kinetickými a termodynamickými koncepty, jak je to dále zde vymezeno, a musí být toto vysvětlení publikováno ve velmi respektovaném recenzovaném vědeckém časopisu/časopisech.106
Do současnosti nebyla žádná odměna vyplacena, a čím víc toho víme o minimálních požadavcích pro život, tím méně pravděpodobné se zdá, že bude nalezeno nějaké alespoň vzdáleně přijatelné materialistické vysvětlení vzniku života. Tento problém je od řešení dál než kdykoli předtím.107
13.1. A důkazy stále v nedohlednu…
Výše uvedené pojednání rozebralo běžné chemické evoluční představy o pravěké chemické polévce, ale zdá se, že jen málo lidí si uvědomuje, že o existenci nějaké takové dávné chemické polévky není ani nejmenší důkaz. Předpokládalo se, že takováto polévka by byla zdrojem základních aminokyselin a nukleotidů, obsahujících dusík. Pokud by existovala, museli by evoluční geologové najít v horninách (o kterých tvrdí, že jsou velmi staré) sedimenty bohaté na dusík. Ale ani v těchto organických materiálech není skoro žádný dusík – jen kolem 0,015 %. Dva geochemici poukazují na to, že:
- Pokud skutečně někdy existovala prebiotická polévka, dalo by se očekávat, že alespoň někde na této planetě nalezneme buď masivní sedimenty obsahující ohromné množství různých organických dusíkatých sloučenin, aminokyselin, purinů, pyrimidinů, atd., nebo dusíkatého koksu [grafitu podobného materiálu obsahujícího dusík]. Ve skutečnosti nebyla žádná taková hmota nikde na Zemi nalezena.108
13.2. Záchrana přichází z vesmíru
Snad aby ještě zdůraznili zoufalství všech těch teorií o chemické evoluci, někteří badatelé tvrdí, že život začal v kosmickém prostoru. Tato myšlenka se nazývá panspermie, z řekého πάν (pan, vše) a σπέρμα (sperma, semeno), čili, že semena života jsou všude ve vesmíru.
Klasický myšlenkový proud panspermických teorií má za to, že semena přišla na zem přirozeně, asi na kometách. Jenže tohle skutečně nic neřeší a jen to daný problém vrhá někam jinam. Problémy s chemií a informací se tím nezmění. Kromě nepravděpodobného přežití miliónů let putování přes chladný, vzduchoprázdný prostor a cesty k našemu slunci s jeho sterilizujícími účinky radiace, nedávný výzkum ukazuje, že mikrobi by nepřežili extrémní teplotu způsobenou třením se zemskou atmosférou – to je ono teplo, které způsobuje efekt ´meteoru´.109
Nová verze této teorie se nazývá řízená panspermie, která tvrdí, že život na tuto zem nasadili cizinci. Zastánci této teorie se opírají o jednoho ze spoluobjevitelů dvoušroubovice DNA, Francise Cricka (1916-2004), a také o Leslie Orgela.110 Zvláště Cricka deptala frustrace z teorií o chemické evoluci:
- Čestný člověk, vyzbrojený veškerým dnes dostupným poznáním, může říct jen to, že se původ života dnes v jistém smyslu jeví téměř jako zázrak, neboť aby k tomu došlo, muselo by být uspokojeno velké množství podmínek. Pokaždé, když píšu článek o vzniku života, zapřísahám se, že už nikdy žádný nenapíšu, protože existuje příliš mnoho spekulací, které vycházejí z příliš malého množství faktů.111
Ještě jednou – toto jen posouvá problém o krok zpět; oni musejí věřit, že tito hypotetičtí cizinci nejprve vznikli chemickou evolucí.112 Člověk žasne, jak může někdo napadat Stvoření jako ´nevědecké´ jen proto, že předpokládá nepozorovatelného Stvořitele, ale za ´vědu´ považuje řízenou panspermii navzdory ad hoc teoriím o nepozorovatelných cizincích! Všimněte si také, že panspermie se opírá o argument založený na ´mezerách v evoluci´ a řízená panspermie se odvolává na projekt (design).
14. Závěrem
Vznik života z neživých chemikálií je článkem slepé víry, nikoli vědy – a toto je pravda už od dob Darwina. Hlavním problémem toho mnohého teoretizování kolem chemické evoluce je to, že teoretikové považují život spíše za shluk chemikálií než za stroj na zpracování informací, a nikdy nezodpoví otázku „Jak si molekulární hardware napsal svůj vlastní software“.
Přírodní výběr nemůže vysvětlit vznik prvního života. Ke zpracování této informace je třeba strojů. Ale instrukce ke stavbě těchto strojů jsou obsaženy v této informaci. A tak problémy kuře-vejce jsou i nadále nezměrné. Tyto stroje potřebují energii a tu dodává zařízení ATP syntáza. Ale motor ATP syntázy nemůže být postaven bez instrukcí a bez čtecího mechanizmu, což je obojí zakódováno v DNA, a montáž tohoto mechanizmu vyžaduje ATP k jeho sestavení. Tohle pak představuje problém vajíčko-larva-kobylka.
Další enzymy jsou pak podstatné pro urychlení životně důležitých reakcí, které by bez nich trvaly milióny let. Jenže samotné enzymy jsou destruktivní, například izolovaná ATP syntáza zničí ATP, nevytvoří jej.
Proteiny se nemohou reprodukovat a v přírodě se rozpadnou. Dlouhé nukleotidy a polysacharidy se také rozpadnou hydrolýzou (ve skutečnosti se ani nikdy nevytvoří). RNA je křehký enzym a je dokonce méně stabilní než DNA. Některé stavební kameny RNA se během pokusů ´simulujících´ chemickou evoluci nevytvořily. Jiné jsou zředěné, kontaminované a/nebo nestabilní. A co víc, nejsou to ´stejně točivé´ formy, jaké vyžaduje život.
A potom, v ´nejranějších´ horninách neexistují žádné důkazy o dávných a nezbytných etapách chemických předstupňů života.
Shrnuto a podtrženo, zákonitosti fyziky, chemie a pravděpodobnosti jsou zcela v rozporu s hypotézami o abiotickém původu života. Nejsou to kreacionisté, kdo se zde odvolává na argument boha mezer! Ve skutečnosti tvrdíme, že jediným rozumným závěrem z toho, co jsme se za ta staletí experimentální vědy naučili, je existence inteligentního Boha stvořitele, který je mimo vesmír a je jeho příčinou.
15. Články na podobné téma v češtině:
- Vznik života
- Vývoj názorů na vznik života až po biblický kreacionismus
- 16 kroků k vygenerování rozvinutého života
- Vznik života-videorozhovor
- Tornáda, led a buňky: Jak termodynamika vzdoruje scénářům o vzniku života
- Evoluce: ta není jen o biologii
- Pět ateistických zázraků
- Původ života: svět RNA?
- Evolucionistické fantazmagorie tryskají bezuzdně z darwinistických lebek
16. Odkazy:
1. Anonymous, Last universal common ancestor more complex than previously thought, ScienceDaily, 3 October 2012; sciencedaily.com.
2. See Lamont, A., Louis Pasteur (1822–1895), Creation 14(1):16–19, 1991; creation.com/louis-pasteur.
3. Darwin, C., On the Origin of Species, 1st ed., John Murray, London, 1859.
4. Letter to Joseph Hooker, 1863, in Darwin, F. (ed), The Life and Letters of Charles Darwin, vol. 3, p. 18, John Murray, London, 1887; accessed via darwin-online.org.uk.
5. Darwin, F. (ed.), The Life and Letters of Charles Darwin, Vol. II, D. Appleton & Co., New York, pp. 202–203, 1911.
6. For a detailed refutation of many of Dawkins’ most popular arguments, see Sarfati, J.,The Greatest Hoax on Earth?, Creation Book Publishers, Powder Springs, GA, USA, 2010, available through creation.com.
7. Flew, A. interviewed by Habermas, G., My pilgrimage from atheism to theism, Philosophia Christi, Winter 2004; biola.edu.
8. Flew, A., with Varghese, R., There is a God, Harper Collins, New York, 2007. See also review by Cosner, L., J. Creation 22 (3):21–24, 2008; creation.com/flew.
9. Johnson, P.E., Response to Gould, Origins Research 15(1):10–11, 1993; www.arn.org. See additional documentation by O’Leary, D., Darwinist Theodosius Dobzhansky was not an orthodox Christian believer! post-darwinist.blogspot.com, 8 September 2006.
10. Dobzhansky, T.G., quoted in Schramm, G., Discussion of Synthesis of Nucleosides and Polynucleotides with Metaphoric Esters, in: Fox, S.W. (ed)., The Origins of Prebiological Systems and of Their Molecular Matrices, pp. 309–310, 1963, Academic Press, New York, 1965.
11. Slack, G., What neo-creationists get right, The Scientist, 20 June 2008; the-scientist.com.
12. Kerkut, G.A., Implications of Evolution, Pergamon, Oxford, p. 157, 1960.
13. John Burdon Sanderson Haldane FRS (1892–1964) is best known as a mathematical population geneticist, but he also worked on enzymes at Cambridge, formulating the Briggs–Haldane Law of enzyme kinetics in 1925. Also a staunch atheist and communist, he proposed a naturalistic origin of life in The Origin of Life, see Rationalist Annual, p.148, 1928.
14. Dickerson, R.E., Chemical evolution and the origin of life, Scientific American 239 (3):62–102, September 1978.
15. Pleasant, L.G. and Ponnamperuma, C., Chemical evolution and the origin of life, Origins of Life and Evolution of Biospheres 10(1): 69–85, 1980.
16. Haeckel, E., The History of Creation, translated by Prof. E. Ray Lankester, M.A., F.R.S, 3rd ed., Vol. 1, p. 184, Kegan Paul, Trench & Co., London, 1883. See also Grigg, R., Ernst Haeckel: Evangelist for evolution and apostle of deceit, Creation 18(2):33–36, 1996; creation.com/Haeckel.
17. Bradbury, S., The Microscope Past and Present, Pergamon Press, 1968; van Niekerk, E., Countering Revisionism, J. Creation 27(1): 78–84, 2013; creation.com/haeckel-fraud-proven-part-2.
18. Dawkins, R., The Blind Watchmaker, W.W. Norton, New York, p. 115, 1986.
19. Dawkins, R.,The Greatest Show on Earth, Free Press, New York, p. 405, 2009.
20. Polanyi, M., Life’s irreducible structure, Science 160:1308, 1968.
21. Orgel, L., The Origins of Life, John Wiley, New York, p. 189, 1973.
22. Davies, P., Life force, New Scientist 163(2204):27–30, 1999.
23. Williams, A., Life’s irreducible structure, J. Creation 21(1):109–115, 2007; creation.com/autopoiesis.
24. Popper, K.R., Scientific Reduction and the Essential Incompleteness of all Science, in Ayala, F. and Dobzhansky, T. (eds.), Studies in the Philosophy of Biology, University of California Press, Berkeley, CA, USA, p. 270, 1974.
25. Dawkins, R., ref. 19, p. 420.
26. Revyakin, A. et al., Abortive initiation and productive initiation by RNA Polymerase involve DNA scrunching, Science 314(5802):1139–1143, 2006; Kapanidis, A.N. et al., Initial transcription by RNA polymerase proceeds through a DNA-scrunching mechanism, Science 314(5802):1144–1147, 2006; see also Nanotech tools yield DNA transcription breakthrough; physorg.com, 16 November 2006.
27. Roberts, J.W., RNA Polymerase, a scrunching machine, Science 314(5802):1139–1143, 2006.
28. Garrett, R., Mechanics of the ribosome, Nature 400(6747):811–812, 1999.
29. Truman, R. and Borger, P., Genetic code optimisation: Part 1, J. Creation 21(2):90–100, 2007; creation.com/gencode.
30. Truman, R. and Borger, P., ref. 29.
31. Sarfati, J., Decoding and editing designs: double-sieve enzymes, J. Creation 13(1):5–7, 1999; creation.com/doublesieve; Karlson, P., (tr. Doering,71- C.H.), Introduction to Modern Biochemistry, 4th ed., Academic Press, London and New York, pp. 113, 145–146, 1975.
32. Nureki, O. et al., Enzyme structure with two catalytic sites for double-sieve selection of substrate, Science 280(5363):578–582, 1998.
33. Sarfati, J., DNA: marvelous message or mostly mess? Creation 25(2):26–31, March 2003; See the animation ‘DNA translation’ at creation.com/message.
34. Aw, S.E., The Origin of Life: A critique of current scientific models, J. Creation 10(3):300–314, 1996; creation.com/origin-of-life-critique.
35. Bergman, J., ATP: The perfect energy currency for the cell, Creation Res. Soc. Q. 36(1):2–10, 1999; creationresearch.org.
36. Hiroyuki Noji et al., Direct observation of the rotation of F1-ATPase, Nature 386(6622):299–302, 1997.
37. Note that it is a subscript letter O not the number zero, for historical reasons: from ‘oligomycin binding fraction’. The antibiotic oligomycin specifically blocks the proton channel on the FO half of bacterial ATP synthase, with deadly effect.
38. Sarfati, J., Design in living organizms (motors), J. Creation 12(1):3–5, 1998; creation.com/motor. See also Thomas, B., ATP synthase, Creation 31(4):21–23, 2009; creation.com/atp-synthase. More details can be found in Sarfati, J., By Design, chapter 10, “Motors”, available through creation.com.
39. Davies, K.M.et al., Macromolecular organization of ATP synthase and complex I in whole mitochondria, Proc. Nat. Acad. Sci. USA 108(34):14121–14126, 2011.
40. Toyabea, S.et al., Thermodynamic efficiency and mechanochemical coupling of F1-ATPase, Proc. Nat. Acad. Sci. USA 108(44):17951–17956,2011.
41. Block, S.M., Real engines of creation, Nature 386(6622): 217–219, 1997 (perspective on Hiroyuki Noji et al., ref. 32).
42. Catalysts do not affect the equilibrium, but only the rate at which equilibrium is reached. They work by lowering the activation energy, which means decreasing the energy of a transitional state or reaction intermediate. See diagram and explanation in Wieland, C. and Sarfati, J., Dino proteins and blood vessels: are they a big deal? 9 May 2009; creation.com/dino-proteins.
43. Miller, B.G. et al., Anatomy of a proficient enzyme, Proc. Nat. Acad. Sci. USA 97(5):2011–2016, 2000.
44. Meaning the reaction would take 78 million years to get half-way to completion. Using the half-life of a reaction is a common way to designate reaction rates in chemistry.
45. Cited in Lang, L.H., Without enzyme catalyst, slowest known biological reaction takes 1 trillion years: study, UNC School of Medicine 262:30, 2003; unc.edu. See also Wolfenden, R. and Snider, M.J., The depth of chemical time and the power of enzymes as catalysts, Acc. Chem. Res. 34:938–994, 2001.
46. This was orotidine 5΄-monophosphate decarboxylase, responsible for de novo synthesis of uridine 5΄-phosphate, an essential precursor of RNA and DNA, by decarboxylating orotidine 5΄-monophosphate (OMP).
47. More detail can be found in Sarfati, J., World record enzymes, J. Creation 19(2):13–14, 2005; creation.com/enzymes and in Sarfati, J., By Design ch. 11, available through creation.com.
48. Lad, C., Williams, N.H., and Wolfenden, R., The rate of hydrolysis of phosphomonoester dianions and the exceptional catalytic proficiencies of protein and inositol phosphatases, Proc. Nat. Acad. Sci. USA 100(10):5607–5610, 2003.
49. Lang, L.H., ref. 45.
50. Yet a virus has a powerful nano-motor to wind up DNA for packaging; Sarfati, J., Virus has powerful mini-motor to pack up its DNA, J. Creation 22(1):15–16, 2008; creation.com/virusmotor; Fuller, D.N. et al., Single phage T4 DNA packaging motors exhibit large force generation, high velocity, and dynamic variability, Proc. Nat. Acad. Sci. USA 104(43):16868–16873, 2007. uote>
51. Fraser, C.M. et al., The minimal gene complement of Mycoplasma genitalium, Science 270(5235):397–403, 1995; perspective by Goffeau, A., Life with 482 genes, Science 270(5235):445–446. Other reports have different numbers, but all within the same ball park.
52. Wood, T.C., Genome decay in the Mycoplasmas, Impact 340, 2001; icr.org.
53. Wells, W., Taking life to bits, New Scientist 155(2095):30–33, 1997.
54. Glass, J.I. et al., Essential genes of a minimal bacterium, Proc. Nat. Acad. Sci. USA 103(2):425–430, 2006.
55. Lane, N., Was our oldest ancestor a proton-powered rock? New Scientist 204(2730):38–42, 2009.
56.Seufferheld, M. et al., Evolution of vacuolar proton pyrophosphatase domains and volutin granules: clues into the early evolutionary origin of the acidocalcisomes, Biology Direct 6:50, 2011.
57. University of Illinois at Urbana-Champaign, ref. 1.
58. Yockey, H.P., A Calculation of the probability of spontaneous biogenesis by information theory, J. Theor. Biol. 67:377–398, 1977.
59. Truman, R., The ubiquitin protein: chance or design? J. Creation 19(3):116–127, 2005; creation.com/ubiquitin. Aaron Ciechanover, Avram Hershko and Irwin Rose won the Nobel Prize in Chemistry in 2004 “for the discovery of ubiquitin-mediated protein degradation”; nobelprize.org.
60. Actually, for very low probability p of success, and d = 1/p, a good rule of thumb is: to have a 95% chance of at least one success, we need 3d trials. In this example, p is about 10–5000, so d = 105000, so we would need about 3 × 105000 trials for a 95% chance of obtaining all the needed enzymes. Personal communication from Dr Jim Davidson (North Carolina), 2012.
61. Quoted in Major, E.L., Big enough to bury Darwin, Guardian (UK) education supplement, 23 August 2001; creation.com/hoyle-origin-of-life. See also Demme, G. and Sarfati, J., Big-bang critic dies, J. Creation 15(3):6–7, 2001; creation.com/hoyle.
62. After Sarfati, J., Self-replicating enzymes? J. Creation 11(1):4–6, 1997; creation.com/replicating.
63. Dawkins, R., ref. 19, pp. 419-420.
64. Woese, C., The Genetic Code, Harper and Row, New York, 1967.
65. Press Release: The 1989 Nobel Prize in Chemistry, 12 October 1989; nobelprize.org
66. Dawkins, R., ref. 18, p. 421.
67. See also Mills, G.C. and Kenyon, D.H., The RNA world: a critique, Origins and Design 17(1): 9–16, 1996; www.arn.org.
68. Shapiro, R., Prebiotic cytosine synthesis, Proc. Nat. Acad. Sci. USA 96(8):4396–4401, 1999.
69. Larralde, R., Robertson, M.P. and Miller, S. L., Rates of decomposition of ribose and other sugars: implications for chemical evolution, Proc. Nat. Acad. Sci. USA 92:8158–8160, 1995.
70. Sarfati, J., Hydrothermal origin of life?, J. Creation 13(2):5–6, 1999; creation.com/hydrothermal.
71. Levy, M. and Miller, S. L., The stability of the RNA bases, Proc. Nat. Acad. Sci. USA 95(14):7933–7938, 1998.
72. Johnston W.K.et al., RNA-catalyzed RNA polymerization, Science 292(5520):131925, 2001.
73. Sarfati, J., Origin of life: the polymerization problem, J. Creation 12(3):281–284, 1998; creation.com/polymer.
74. Sarfati, J., Origin of life: the chirality problem, J. Creation 12(3):263–266, 1998; creation.com/chirality.
75. Joyce, G.F. et al., Chiral selection in poly(C)-directed synthesis of oligo(G), Nature 310:602–604, 1984.
76. Johnston, ref. 74, admints that their ribozyme’s copying accuracy is “still lower than the ≥0.996 fidelity seen with viral polymerases that replicate RNA by using RNA templates,and it is much lower than that seen for polymerases that replicate DNA.”
77. Further chemical problems are found in Cairns-Smith, A.G., Genetic Takeover: And the Mineral Origins of Life, Cambridge University Press, 1982; see extract at creation.com/rna.
78. Schirber, M., A salt-free primordial soup? Astrobiology Magazine, 19 January 2012; astrobio.net.
79. Joyce, G.F., RNA evolution and the origins of life, Nature 338:217–224, 1989.
80. Harold C. Urey—Biography, nobelprize.org.
81. Miller, S. L., A production of amino acids under possible primitive earth conditions, Science 117:528–529, p. 528, 1953; Miller, S. L., Production of some organic compounds under possible primitive earth conditions, J. Amer. Chem. Soc. 77:2351–2361, 1955.
82. Yockey argued in Nature (415(6874):833, 2002) that Stanley Miller wasn’t the first. There were earlier experiments of Walther Löb (1913), Oskar Baudisch (1913), Edward Bailey (1922) and Harold Urey (1928, 1929). Yockey suggested that Miller merely augmented these previous experiments with modern separation and detection techniques such as two-dimensional paper chromatography. Coincidentally, the significance of these techniques was emphasized by my organic chemistry professor. A reply by Jeffrey Bada and Antonia Lazcano (Nature 416(6880):475, 2002) defended the significance of Miller’s experiments for chemical evolution, while Löb showed no interest in this.
83. Bergman, J., Why the Miller–Urey research argues thagainst abiogenesis, J. Creation 18(2):74–84, 2002; creation.com/urey.
84. Allentoft, M.E. et al., The half-life of DNA in bone: measuring decay kinetics in 158 dated fossils, Proc. Royal Society B 279(1748):4724–4733, 2012.
85. Newly discovered DNA repair mechanism, Science News, sciencedaily.com, 5 October 2010.
86. Shapiro, J.A., A third way,Boston Review, p. 2, February/March 1997; Sarfati, J., New DNA repair enzyme discovered, creation.com/dna-repair, 13 January 2010.
87. Sarfati, J., ref. 74.
88. E.g. the amino group (–NH2) in the amino acid reacts readily with the carbonyl group (O=C<) in the sugar, releasing a water molecule (H2O) to form an imine (HN=C<), which is useless for life. See Bergman, ref. 83.
89. If such a soup produced all the nitrogenous compounds required for life, why is there no trace of them in the ‘earliest’ rocks? Cf. Brooks, J. and Shaw, G., Origins and Development of Living Systems, Academic Press, London, UK and New York, 1973.
90. Sarfati, J., ref. 74.
91. Nielsen-Marsh, C., Biomolecules in fossil remains: Multidisciplinary approach to endurance, The Biochemist, pp. 12–14, June 2002. See also Doyle, S., The real ‘Jurassic Park’, Creation 30(3):12–15, 2008; creation.com/realjurassic, and Thomas, B., Original animal protein in fossils, Creation 35(1):14–16, 2013; creation.com/ancient-protein.
92. This is the simple Arrhenius rate equation k = A exp(-Ea/RT), where k is the rate constant, A is a temperature-independent constant (often called the frequency factor), exp is the exponential function, Ea is the activation energy, R is the universal gas constant, and T is the absolute temperature.
93. Sarfati, J., ref 6., pp. 204-208.
94. Schweitzer, M.H. et al., Molecular analyses of dinosaur osteocytes support the presence of endogenous molecules, Bone 52(1):414–423, 2013; see also Sarfati, J., DNA and bone cells found in dinosaur bone, J. Creation 27(1):10–12, 2013; creation.com/dino-dna.
95. The evolutionist Cairns-Smith has raised the same objections against the typical ‚origin of life‘ simulation experiments in ref. 79.
96. Thaxton, C.B., Bradley, W.L., and Olsen, R.L., The Mystery of Life’s Origin, ch. 6, Philosophical Library Inc., New York, 1984.
97. Williams, A., Life’s irreducible structure—Part 1: autopoiesis, J. Creation 21(2):116–122, 2007; creation.com/autopoiesis.
98. De Duve, C., Singularities: Landmarks on the Pathways of Life, Cambridge University Press, Cambridge, 2005. See review by Williams, A., Great minds on the origin of life, J. Creation 21(1):38–42, 2007; creation.com/singularities.
99. Conway Morris, S., Life’s Solution: Inevitable humans in a lonely universe, Cambridge University Press, Chs 3–4, 2003. See review by ReMine, W., Evidence for Message Theory, J. Creation 20(2):29–35, 2006; creation.com/lifes-solution.
100. Cairns-Smith, A.G., Genetic Takeover and the Mineral Origins of Life, Cambridge University Press, New York, 1982.
101. Photosynthesis is another irreducibly complex system, storing up the energy of four photons to split the strongly bound water molecule. See Sarfati, J., Green power (photosynthesis): God’s solar power plants amaze chemists, J. Creation 19(1):14–15, 2005; creation.com/greenpower.
102. Hulett, H.R., Limitations on prebiological synthesis, J. Theor. Biol. 24:56–72, 1969.
103. UV-B penetrates 65 metres deep in clear Antarctic waters , according to Gieskes, W.C. and Kraay, G.W., Transmission of ultraviolet light in the Weddell Sea: Report on the first measurements made in Antarctic, Biomass Newsl. 12:12–14, 1990.
104. Yockey, H.P., ref. 58, p. 379.
105. Yockey, H.P., ref. 58, p. 396.
106. The Origin-of-Life Prize; www.us.net/life.
107. Smith, Calvin, Who wants to be a millionaire, creation.com/lifeprize, 15 Aug 2007.
108. Brooks, J., and Shaw, G., Origins and Development of Living Systems, Academic Press, London and New York, 1973.
109. Meteorite experiment deals blow to bugs from space theory, Physorg.com, 25 September 2008; Sarfati, J., Panspermia theory burned to a crisp: bacteria couldn’t survive on meteorite, 10 October 2008; creation.com/panspermia.
110. Crick, F. and Orgel, L.E., Directed Panspermia, Icarus 19:341–346, 1973.
111. Crick, Life Itself, Its Origin and Nature, pp. 88, 153, Simon and Schuster, 1981.
112. See also Bates, G., Designed by aliens? Creation 25(4):54–55, 2003; creation.com/aliens.