
Daniel Criswell, Ph.D.
molekulární biolog
Z ICR (AB0 Blood and Human Origins) přeložil M. T.
Mnoho lidí ví, jaká je jejich krevní skupina a chápou, že krevní skupiny musí být totožné při poskytování první pomoci. Skupiny systému AB0 jsou nejdůležitějším krevním faktorem v klinických aplikacích vyžadujících transfúze krve. Chápání důležitosti systému AB0 není však omezeno jen na klinické aplikace. S naší novou schopností rychle sekvenovat geny, se systém AB0 osvědčuje také jako cenný nástroj k určování vzorců lidské migrace a původu člověka.
Co určuje krevní skupinu?
Krevní skupiny systému AB0 určuje ukazatel (marker) na povrchu buňky, který identifikuje buňku jako patřící ke „svým“ čili k určitému jedinci. Tyto markery na povrchu buněk jsou charakterizovány proteinem či lipidem, který se navazuje na určité uspořádání cukrů.
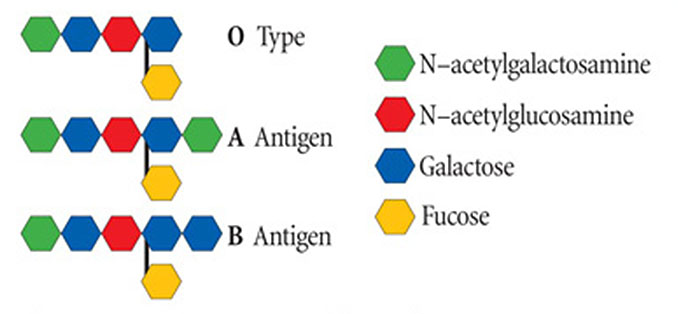
Obrázek 1 – antigenní určení systému AB0. Vidíme uspořádání cukrů, jež určuje každý z typů krvinek A, B, a 0 (1). Antigeny AB0 se liší jen jedním sacharidem na konci antigenu. Na obrázku je pouze uhlovodanová část antigenu. Všimněte si, že všechny jsou stejné, až na to, že skupiny A a B mají cukr navíc: N-acetylgalaktosamin pro A, a galaktosu pro B.
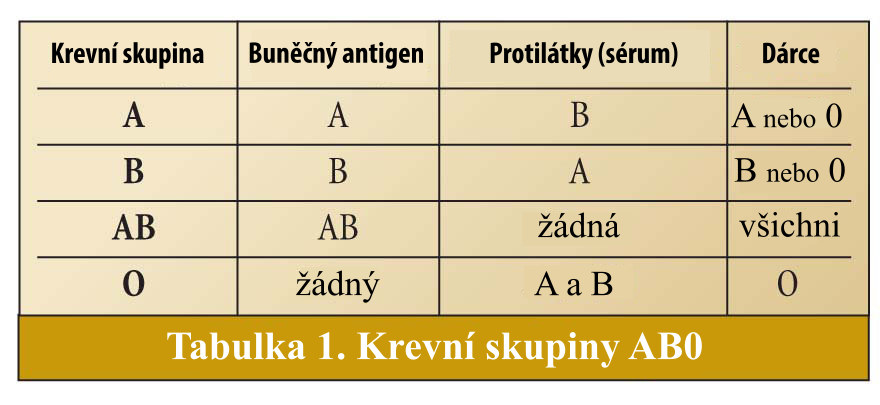
Uspořádání cukrů je součástí antigenu schopného odstartovat imunitní odpověď, která vyprodukuje protilátky k identifikaci a zničení cizích antigenů. Lidé s krevní skupinou A produkují protilátky B, když jsou vystaveni antigenu B, a ti s krevní skupinou B produkují protilátky A, jsou-li vystaveni antigenu A. Krevní skupina AB však protilátky neprodukuje, protože oba antigeny přítomné na buňkách jsou rozpoznány jako „vlastní“. Krevní skupina 0 produkuje protilátky A i B, protože ani antigen A ani B není přítomen na buňkách jedinců skupiny 0 (tabulka 1).
Protilátky A a B patří do „M“ třídy imunoglobinů a jsou vyjádřeny imunoglobinovými geny lymfocytů B při vystavení cizím antigenům. Imunoglobinové geny jsou schopny produkovat v podstatě neomezené množství protilátek složitým procesem editace a selekce (1). V důsledku toho neexistuje specifický gen pro protilátky A či protilátky B děděný s komplementárním antigenem A nebo B.
Gen pro specifikaci antigenů A či B či 0 určuje krevní skupinu. Enzym, glykosyltransferáza, je produktem tohoto genu (2), a rozdíly v sekvenci tohoto enzymu (polymorfizmy) určují, zda enzym naváže N-acetylgalaktosamin (antigen A), galaktosu (antigen B), nebo žádný cukr (obrázek 1). Lidé dědí dva geny pro krevní skupinu; nebo, přesněji řečeno, dvě alely, jednu od každého rodiče.
Tyto alely označujeme jako IA pro skupinu A , IB pro skupinu B a i pro skupinu 0. Obě alely glykosyltransferázy pro antigeny A a B jsou vyjádřeny, pokud jsou zděděny spolu, a produkují oba antigeny a tvoří krevní skupinu AB. Pokud je alela pro krevní skupinu A či B děděna se skupinou 0, jedinec bude mít krevní skupinu buď A nebo B. To není nutně proto, že alela typu 0 je potlačená čili recesivní, ale je to spíše výsledek aktivity glykosyltransferázy A či B, zatímco glykosyltransferáza pro alelu 0 je neaktivní (2). Jedinec skupiny 0 má obě alely pro inaktivní glykosyltransferázu.
Pokud by Adam a Eva byli heterozygoty pro krevní skupinu A respektive B, byli by mohli mít děti, které by měly kteroukoli z krevních skupin AB0.
Heterozygot = jedinec, který ve svém genovém páru má dvě různé alely. Opak: homozygot.
Krevní skupiny a původ člověka
Takže jaké světlo vrhají tato zjištění na původ člověka? Je možné, aby dva lidé ze zprávy o Stvoření (Adam a Eva) či osm lidí v arše Nóachově dali vzniknout všem typům krve AB0 přítomným u lidí dnes? Pokud by Adam a Eva byli heterozygoty pro krevní skupinu A respektive B (jedna alela pro skupinu 0 a jedna alela buď pro typ A či B), byli by mohli mít děti, které by měly kteroukoli z krevních skupin AB0, jak ukazuje obrázek 2. Punnettův čtverec jednoduše předpovídá, jaké by byly možné fenotypy pro děti daného páru. Z mnoha dětí, jež Adam a Eva nejspíš měli, není těžké předvídat všechny typy krve AB0, které zdědili jejich potomci.
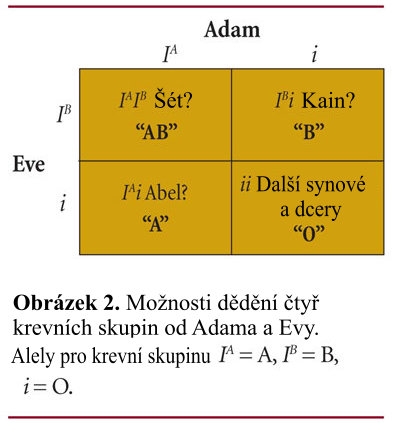
Pokud byli Adam a Eva heterozygoti pro genový lokus krevních skupin AB0, pak četnost alel pro skupinu 0 je 50 procent (2 ze 4 alel), alelová četnost pro typ A je 25 procent (1 ze 4 alel), a alelová četnost pro typ B je 25 procent (obrázek 2). Neexistují-li selektivní tlaky či genetický drift pro tyto alely, pak alelová četnost zůstane táž pro veškeré jejich potomstvo. Celková alelová četnost v Punnettově čtverci je vlastně táž pro děti, jaká asi byla pro Adama a Evu. Tento scénář by platil také pro Nóachovu rodinu a jejich potomky.
Lokus = místo. V genetice místo na chromozomu, kde je lokalizován určitý gen.
Četnosti alel v moderní populaci
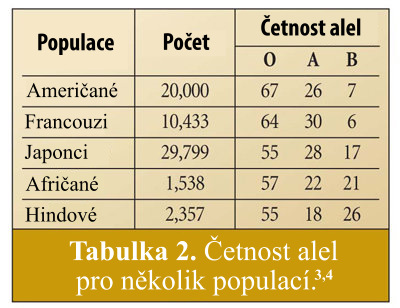
Odrážejí lidské populace dnes tyto alelové četnosti? Odpověď zní ano. Tabulka 2 ukazuje alelové četnosti několika populací. (Všimněte si, že nejde o četnosti krevních skupin). Existuje všeobecný vzestup četnosti alely typu 0, a v mnoha populacích úpadek v typu B alely. Ale jak jsme očekávali, četnosti pro každou alelu se blíží hodnotám, jaké by byly mohly být na počátku historie lidstva či u Nóachovy rodiny. Posun v četnosti (vzestup u typu 0 a úbytek u typu B) může být způsoben migrací lidských skupin, jež měly vyšší či nižší četnost jedné z alel v době migrace. Příčinou může být též náhodný genetický drift, či mutace, jež činí glykosyltransferázu neaktivní – což by se projevilo v krevní skupině 0 z typu A; jde zřejmě o jednu z příčin pro vzestup v četnosti alely 0.
Bohužel, původ alel AB0 se zamotává, zkoumáme-li aktuální gen pro glykosyltransferázu. Existuje více než 180 variací (polymorfizmů) pro gen AB0 uváděných na webové stránce Národního centra pro biotechnologické informace (NCBI) (5), a jeden každý z těchto polymorfizmů může být přiřazen k jedné ze tří alel AB0. Většina z těchto polymorfizmů nemění aktivitu glykosyltransferázy či krevní skupinu, ale může identifikovat etnické skupiny, jež se utvořily poté, co lidé osídlili celou zemi. Mutace a chromozomový crossing-over jsou nejpravděpodobnější příčinou těchto variant (6).
Crossing-over = překřížení odpovídajících částí chromatid analogických chromozomů během meiózy. Tento proces umožňuje vznik nových kombinací vlastností u potomků.
Existují rozdíly DNA čili polymorfizmy, které určují funkci glykosyltransferázy, což vede k různým typům krve AB0. Těchto rozdílů je málo, ale jsou nezanedbatelné. Glykosyltransferáza specifická pro syntézu antigenu A se liší od enzymu specifického pro antigen B jen ve čtyřech aminokyselinových zbytcích (z 354), a existuje několik odlišností v sekvenci DNA v alelách, které kódují specifický enzym pro krevní skupiny A a 0 (2). Ony čtyři rozdíly mezi glykosyltransferázami A a B stačí k tomu, aby enzym označil charakteristický terminální cukr, který odlišuje antigeny A a B. Jediné vymizení DNA v alele specifické pro A ústí do okleštěné verze glykosyltransferázového genového produktu a eliminuje enzymatickou aktivitu a jeho výsledkem je nakonec krevní skupina 0.
Důsledky původu krevní skupiny „0“
Je možno namítat, že jedna ze tří alel zrodila další dvě. Například, původ alely 0 a následně i krevní skupiny 0, je prostě výsledkem vymizení (delece) ústícího do ztráty funkce glykosyltransferázové aktivity pro antigen A. Mutace ústící do ztráty funkce u proteinu, přinejlepším, by byla „téměř neutrální“ mutací, jelikož krevní skupina 0 zdá se nemá žádné zhoubné účinky či selektivní výhodu nad ostatními dvěma typy krvinek.
Protože neutrální či téměř neutrální mutace nemají selektivní výhodu, je prakticky nemožné fixovat tyto mutace do velké populace organizmů (fixace = 100 procent alel v přijatelném čase. Například, pokud by mutace, která dala krevní skupinu 0, byla skutečně o 1 procento výhodnější než typ A, zabralo by to 100 000 generací, než by se tato mutace fixovala v moderní lidské populaci z počáteční populace 10 000 lidí. (7). Čím větší populace v době mutace, tím déle bude trvat, než se zafixuje, a tím menší je pravděpodobnost, že bude vůbec kdy fixována.
Molekulárně evoluční časové stupnice umísťují vznik moderních lidí asi do doby před 200 000 lety, (9), což je časový rámec příliš krátký, aby zvýšil četnost alely 0 na 60 procent všech lidí dnes žijících, z populace 10 000.Biblický časový rámec by byl jistě ještě mnohem kratší pro takovou fixaci. Delece odpovědná za proměnu alely A v alelu 0 se nevyskytuje u šimpanzů, a srovnání sekvencí mezi lidmi a šimpanzi naznačuje, že tato alela je pouze v lidské rodině (10, 11), což dále komplikuje evoluční scénář pro původ krevní skupiny 0. Tenhle scénář by seděl lépe, kdyby alela 0 byla vzácná v dnešní populaci a objevovala se u specifické skupiny lidí. Avšak alela 0 je zdaleka nejrozšířenější globální alelou, což naznačuje, že pokud opravdu vznikla mutační událostí, musela se objevit tehdy, když lidská populace byla extrémně malá, a dříve než se lidé rozdělili do etnických skupin a rozšířili se po zemi.
Je možné dosáhnout současné četnosti alely 0 cestou mutace, pokud k ní došlo v době Noemovy Potopy a byla předávána jedním z členů Noemovy rodiny. Nóach či jeho žena mohli mít alelu 0 a předat ji všem svým synům, nebo mohly alely zmutovat v potomcích jednoho ze synů. Populace lidské rasy v době Potopy a bezprostředně po ní se určitě jeví jako velikost populace, která by umožňovala zmutované alele stát se běžnou tak, jak populace rostla. Se startovní populací pouhých osmi lidí, alela 0 mohla snadno vzrůst v četnosti náhodným genetickým driftem v popotopní populaci, reflektujíc současné úrovně, jež dnes pozorujeme a v souladu s počítačovými simulacemi modelujícími fixaci. (12).
Závěr
Pokud Adam a Eva neměli alely všech tří krevních skupin, muselo dojít k mutaci, která vytvořila alelu 0 pokud byla lidská rasa ještě velmi malá a předtím, než se lidé rozšířili po zemi. Ať už byl původ krevní skupiny 0 v Adamovi a Evě při stvoření či ať už vznikla mutační událostí, k níž došlo krátce před či po Potopě, je to silná podpora pro tvrzení, že všichni dnešní lidé jsou potomky dvou jedinců či malé skupiny lidí, která nakonec osídlila zemi. Oba scénáře jsou v souladu s biblickým modelem lidských počátků.
Odkazy
- Goldsby, R.A. et al. 2000. Kuby Immunology. 4th ed. New York: W.H. Freeman.
- Yamamoto, F. et al. 1990. Molecular genetic basis of the histoblood group ABO system. Nature 345 (6272):229-33.
- Minkoff, E.C. 1983. Evolutionary Biology. Menlo Park, CA: Addison Wesley.
- Sinnot, E.W. et al. 1958. Principles of Genetics. 5th ed. New York: McGraw-Hill Book Company.
- www.ncbi.nlm.nih.gov/gv/rbc/xslcgi.fcgi?
cmd=bgmut/systems_info&system=abo. - Hosseini-Maaf, B. et al. 2003. ABO exon and intron analysis in individuals with the AweakB phenotype reveals a novel O1v-A2 hybrid allele that causes four missense mutations in the A transferase. BMC Genetics 4:17.
- Patterson, C. 1999. Evolution. Ithaca, NY: Comstock Publishing Associates.
- Sanford, J.C. 2005. Genetic Entropy & the Mystery of the Genome. 2nd ed. Lima, NY: Elim.
- Cann, R. L. et al. 1987. Mitochondrial DNA and human evolution. Nature 325 (6099):31-6.
- Kitano, T. et al. 2000. Gene diversity of chimpanzee ABO blood group genes elucidated from intron 6 sequences. The Journal of Heredity 91 (3):211-4.
- Kermarrec, N. et al. 1999. Comparison of allele O sequences of the human and non-human primate ABO system. Immunogenetics 49 (6):517-26.
- Hartl, D.L. and A.G. Clark. 1989. Principles of Population Genetics. 2nd ed. Sunderland, MA: Sinauer Associates Inc.