Evoluce svalů je pro neodarwinizmus velkým problémem: přehled
Jerry Bergman
Z CRSQ roč. 44, č. 1, léto 2007, str. 56 – 60. přeložil M. T. – 09/2007.
Souhrn
Sval má strukturu, jež je nezjednodušitelně složitá a nevykazuje známky postupné evoluce, srovnáváme-li jeho vlastnosti u jednotlivých zvířat. Kosterní svalstvo včely je ve své anatomii a funkci jen málo odlišné od svalstva lidského. Svaly ve všech formách života jsou tak podobné, že to vyžaduje cvičené oko a dobrý mikroskop, abychom určili rozdíly. Zdá se logické uzavřít tuto studii konstatováním, že organizmy každého druhu byly vybaveny vlastními dobře vymyšlenými svalovými tkáněmi.
Úvod
Svaly jsou pro zvířata životně důležité. Užívají se nejen k pohybu ve velkém měřítku, ale i k regulaci mnoha malých životních pochodů jako je pohyb oční bulvy. Lidé potřebují svaly na to, aby zvedli knihu, mrkli okem, učinili krok, pokynuli prstem, hodili míčem či dokonce i k tomu, aby se nadechli. Svalovou tkáň považují vědci za tak důležitou, že jeden badatel, který objevil, jak fungují svalové buňky, řekl, že pracuje v tomto oboru proto, že „se nejniterněji dotýká podstaty života“ (Vogel, 2001, str. 11).
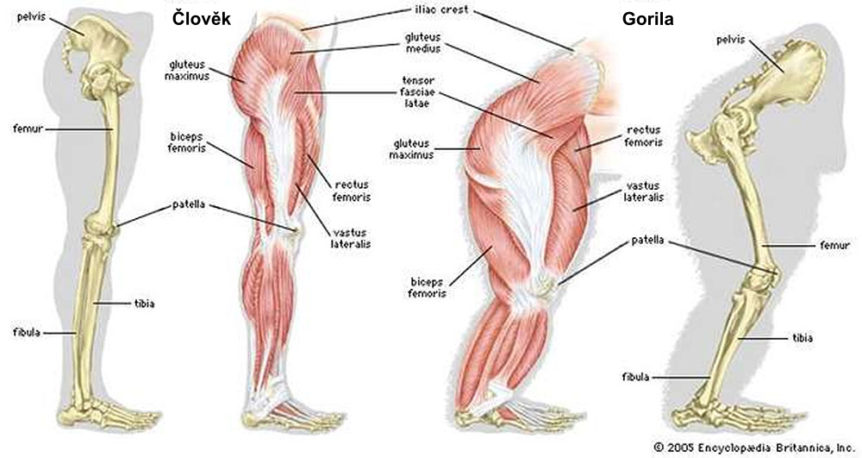
Neodarwinisté předpovídají, že sval u zvířat jako jsou červi, hmyz a savci by měl vykazovat vzestupnou řadu forem a evolučních „výhod“. Nic takového však vědci nezjistili. Studium komparativní anatomie odhalilo, že svaly napříč celou živočišnou říší se liší jen málo v základním plánu i funkci, ať už studujeme včelu nebo člověka. Plní však mnoho různých úkolů jako je lidská chůze, let mouchy, chřestění chřestýše a u olihně vystřelování černých tekutin, aby se ve vodě lépe ukryla. Plán svalu u blech, slonů i lidí je téměř totožný; jeho stavba je u všech tak podobná, že to vyžaduje jak mikroskop tak cvičené oko, abychom rozeznali odlišnosti (viz Vogel, 2001, str. ix, 1).
Svalovina prodávaná jako jídlo se nazývá maso. Potravinářská inspekce musí používat biochemické testy s protilátkami, když se snaží odhalit pokusy podvést zákazníky podvržením levnějšího masa jako je konina za hovězí.
Typy svalů
Čtyři hlavní typy svalů jak u obratlovců tak bezobratlých jsou (Oota a Saitou, 1999):
- rychlé kosterní
- pomalé kosterní
- srdeční
- hladké
Liší se umístěním v organizmu: kosterní svalstvo je připojeno v první řadě ke kostem a občas i k pokožce, srdeční sval tvoří stěny srdce, a svalstvo hladké tvoří stěny dutých orgánů jako jsou cévy, žaludek a střeva.
Kosterní svalstvo a srdeční sval se pod mikroskopem jeví jako příčně pruhované, nikoli hladké (Saladin, 1998). V srdečním svalu je zabudován krokoměr (regulátor vzruchů) k dosažení autorytmicity. Hladké svalstvo používá často též ve svém fungování autorytmicitu. Kosterní svalstvo je kontrolováno vědomě, zatímco srdeční sval a svalstvo hladké jsou do značné míry ovládány nervovými přenašeči vzruchů (neurotransmitery) a hormony. Vogel (2001) konstatuje, že sval je konstruován podle stejného vzoru ve všech živých organizmech:
Tentýž orgán pohání mikroorganizmus tři setiny milimetru dlouhý jako pohání velrybu třicet metrů (čili miliónkrát tolik) dlouhou. Svaly se ovšem mezi sebou skutečně liší – červený sval, bílý sval, vláknitý sval, chytací sval – , nesmíme však pustit ze zřetele implicitní obecnou povahu příčně pruhovaného kosterního svalu. V tom, jak fungují, v rychlosti, s jakou reagují, v produkci síly i výdeji energie se svaly mezi sebou liší méně než je tomu u elektrických motorů, jež pohánějí naše domácí spotřebiče.
Navíc je „architektura daného svalu mimořádně konstantní u jedinců téhož druhu a tytéž základní architektonické zásady platí v celé živočišné říši“ (Lieber a Friden, 2000, str. 1647). Funkci všech svalů lze shrnout takto: nervové podráždění způsobí, že se sval stáhne či se o to pokusí, třeba aby něco podržel. Dokonce i svalové mitochondrie jsou si velmi podobné u bezobratlých i obratlovců (Henwood, 1992). Tato podobnost odporuje evoluční teorii neodarwinistů.
Anatomie a fyziologie svalu
Asi polovinu váhy člověka tvoří svalová hmota. Pět základních funkcí svalové soustavy těla je: (1) umožňovat pohyb těla (jako třeba chůzi), (2) stabilizovat polohu těla (jako třeba stání či sezení), (3) umožňovat pohyb látek v těle (srdce pumpuje krev, stahy hladkého svalstva umožňují pohyb potravy zažívacím traktem), (4) regulovat objem orgánu (jako třeba umožňovat pohyb obsahu dutým orgánem), a (5) fungovat v získávání tepla (svalové stahy vyrábějí až 85 procent veškerého tělesného tepla). Nazýván „elektrický motor“, sval je „pozoruhodným motorem: jemným, dobře namazaným a pružným“ (Vogel, 2001, str. 18).
Veškerá svalová tkáň má pět charakteristických rysů: (1) dráždivost, tj. vlastnost odpovídat na podněty vydáváním elektrických signálů zvaných akční potenciály, jež vedou k pohybu; (2) vodivost, tj. schopnost buňky šířit tyto akční potenciály; (3) stažitelnost, tj. schopnost svalů zkracovat se a nabývat na objemu, což umožňuje pohyb a vykonávání práce; (4) roztažitelnost, tj. schopnost svalu natáhnout se, aniž by se tkáň poškodila; a (5) elasticitu, tj. rys umožňující svalu vrátit se ke svému původnímu tvaru po stahu či natažení (Seeley et al., 2003).
Kosterní svalstvo je dobře zásobováno krví (cévami), aby mělo dostatek živin k vykonávání svých funkcí. Krev je třeba k přepravě živin, jež svalové buňky potřebují k výrobě ATP, k opravě svalových buněk, a také k tomu, aby odnášela odpadní látky vznikající při mnoha chemických reakcích prováděných svalem. Vztahy jako jsou tyto mezi svaly a krevním oběhem ilustrují, jak je samotné tělo, tělo obratlovce i bezobratlého, nezjednodušitelně složité. Každá z mnoha částí těla musí být přítomna a v dobré kondici, aby bylo tělo výkonné. Tenhle typ „nezjednodušitelně složitého“ systému je pádným důkazem nadpřirozeného inteligentního plánu.
Poznámka překladatele: ATP = adenosintrifosfát je energeticky bohatá látka (makroergní fosfát), která je schopna uvolnit energii pro potřeby buňky a do níž se energie rovněž ukládá (AdenosinTriPhosphate).
Nezjednodušitelná složitost – neredukovatelná složitost, angl. irreducible complexity.
Nervové ovládání
Svaly jsou ovládány elektrochemickými signály. Nervový signál putuje cestou vycházející z nervosvalového spojení zvaného synapse, termín znamenající „spojení“. Synapse funguje jako přepínač a je obvykle umístěna v oblasti obsahující malou jamku (synaptickou štěrbinu), jež odděluje dvě nervové buňky. K překlenutí této jamky je třeba neurotransmiteru, který putuje od nervu ke svalu. Na nervosvalovém spojení se synapse mezi neuronem a svalovými vlákny (vzdálenou částí zakončení axonu) rozšiřuje v synaptický koncový hrbolek (bulbus) obsahující množství váčků uzavřených membránou zvaných synaptické váčky. Evoluční původ svalové kontroly takovými elektrochemickými signály zůstává nevysvětlen i po desetiletích výzkumu.
Synaptické váčky obsahují molekuly neurotransmiteru (přenašeče vzruchů). Nervosvalové spojení užívá jako neurotransmiter acetylcholin. Na druhé straně synaptické štěrbiny je motorová koncová destička, jež obsahuje okolo 30 až 40 miliónů acetylcholinových receptorů. Když nervový impuls dojde ke koncovému bulbu synapse, vyvolá uvolnění acetylcholinu, jenž pronikne do synaptické štěrbiny a naváže se na receptor, což vyvolá příliv sodíkových iontů.
Ionty změní klidový potenciál membrány a uvolní svalový akční potenciál, jenž putuje plazmatickou membránou svalové buňky a způsobí stažení svalu (Seeley et al., 2003). Složitost a také prostota tohoto sledu událostí zůstávají v neodarwinizmu nevysvětleny.
Motorické nervy dodají podnět, jenž způsobí stažení svalových buněčných vláken. Neuron a sada asi 150 svalových vláken, jež neuron stimuluje, se nazývají motorická jednotka. Všechny svaly v motorické jednotce se stahují a uvolňují zároveň.
Aby mohly fungovat, potřebují svaly mnoho podpůrných struktur – jejichž původ nelze neodarwinisticky vysvětlit. Sval sám je „polotuhým gelem“, jenž potřebuje mohutné vrstvy pojivové tkáně, aby mohl fungovat (Vogel, 2001, str. 11). Jako příklad uveďme fascie (z latinského fascia znamenající „obvaz“). Jsou to vrstvy vláknité pojivové tkáně, jež obklopují sval.
Nejprve jsou to povrchové fascie, jež jsou bezprostředně pod kůží a zajišťují mechanickou ochranu proti úrazu a oporu pro nervy a krevní cévy, které vstupují do svalů a vycházejí ze svalů. Druhý typ, fascie hloubkové, jsou hustou nepravidelnou pojivovou tkání, která obklopuje svaly, vyděluje je do skupin podle určitých funkcí, umožňuje jim volný pohyb a je oporou pro nervy, krevní cévy a cévy lymfatické (Saladin, 1998).
Tenda – šlachy (z tendere – „napínat“) jsou provazce husté pojivové tkáně, jež upínají sval na kostní periosteum. Aponeurosis (apo, „z“, a neuron, „šlacha“) je vláknitá plachtovitá membrána připomínající rozloženou šlachu. Je to vrstva pojivové tkáně, která se prostírá jako široká plochá vrstva přes sval a upíná ho na obal kostního svalu či na kůži.
Struktura kosterního svalu
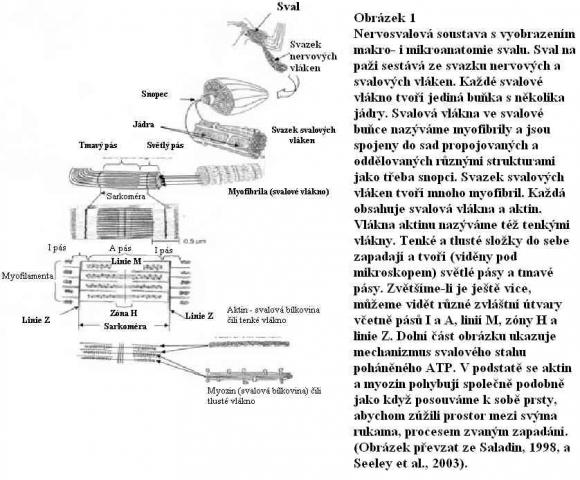
Mikroskopické zkoumání kosterního svalu odhalí stovky či tisíce dlouhých buněk zvaných svalová vlákna čili myofibry, jež jsou uspořádány paralelně jedno k druhému. V průměru mají od 10 do 100 mikrometrů. Plazmatická membrána svalové buňky je sarkolema (sarko znamená „tělo“ a lema znamená „pochva“). Obklopuje cytoplazmu svalového vlákna zvanou sarkoplazma.
Během vývoje embrya vzniká jediný sval ze spojení mnoha menších buněk zvaných myoblasty (zárodečné svalové buňky). Z tohoto důvodu má každé svalové vlákno mnoho jader umístěných po obvodu buňky, pěkně mimo cestu stahových prvků. Mitochondrie jsou okysličovací organely, které spočívají v řadách uvnitř svalových vláken, což je příhodné uspořádání umožňující svalovým proteinům využívat velké množství adenosintrifosfátu (ATP) nutného pro stah
Velmi důležité jsou myofibrily (stahové prvky kosterního svalu), jež obsahují jak tenká, tak tlustá vlákna (Seeley et al., 2003). Tyto buněčné anatomické podrobnosti podporují teorii o původu svalových buněk podle plánu, nikoli postupnou evolucí po krocích. Jedním důvodem pro toto tvrzení je fakt, že sval nebude fungovat, dokud nejsou přítomny všechny jeho části vyobrazené v diagramu. Bez tenkého vlákna, tlustých vláken, jádra, myofibrily a mnoha dalších částí fungovat nebude.
Stah svalu je výsledkem činnosti soustavy do sebe zapadajících a posuvných vláken. Ačkoli nám přesný mechanizmus nutný ke stahu svalu uniká, je známo, že je velmi složitý a vyžaduje množství různých proteinů – alespoň šest typů včetně titanu je účastno v pouhém posunovacím mechanizmu (viz Vogel, 2001, str. 20). Myozinové hlavice táhnou tenká vlákna a posunují je k zóně H umístěné ve středu vláken tlustých. Dojde-li k maximálnímu stahu, pak se tenká vlákna překrývají a způsobují zkrácení svalu.
Vzestup koncentrace iontů vápníku v sarkoplazmě vyvolá pohyb vláken svalu a naopak snížení této koncentrace pohyb zastaví. Aktivní dopravní pumpy dopravují vápník ze sarkoplazmy do organely sarkoplazmatického retikula. Vápník uvolněný ze sarkoplazmatického retikula se spojí s troponinem a způsobí změnu jeho konfigurace, což odstraňuje troponino – tropomyozinový komplex z míst vážících myozin na aktinu.
Tropomyozin = svalový protein tvořící s troponinem komplex, který v klidovém stavu brání reakci aktinu s myozinem. Je součástí aktinového filamenta – vlákna. (pozn. překl.)
Disky Z jsou koncové ploténky (ukončení motorického nervu), které spojují sousední svalové buňky. Vogel (2001) je nazval „komplikované struktury složené z nejméně čtyř dalších druhů proteinů“. Fosfátová sloučenina ATP dodávající velké množství energie zajišťuje většinu energie potřebné pro svalový stah. Když sval odpočívá, váže se ATP na specifická místa na křížových můstcích myozinu umístěných na myozinových výběžcích. Na hlavici myozinu je přítomna část, jež působí jako ATPáza, enzym užívající hydrolýzu ke štěpení ATP na ADP plus fosfátovou skupinu.
ADP = adenosindifosfát. Látka vznikající z ATP odštěpením jedné fosfátové skupiny vázané makroergní vazbou enzymem ATPázou. Při dostatku energie se opět mění na ATP. (pozn. překl.).
Výsledkem je přenos energie z ATP na myozinovou hlavici vybuzující myozinové křížové můstky. Když hladina vápníku v sarkoplazmě stoupne na určitou úroveň, odsune se tropomyozin ze své blokující polohy a způsobí, že se aktivované myozinové hlavice spontánně naváží na místa vážící myozin na aktinu. To opět způsobí náraz stahu síly, což přiměje myozinové hlavice natočit se ke středu sarkoméry, podobně jako se pohybují vesla člunu při veslování. Když se myozinové hlavice otočí, uvolní tím ADP k recirkulaci. Vzájemné vztahy částí přítomných v tomto systému jsou dalším příkladem nezjednodušitelné složitosti, jež vyžaduje plán.
Svalová vlákna se po stahu uvolní, protože acetylcholin je rychle odbourán acetylcholinesterázou. Tak končí vytváření svalového akčního potenciálu a dojde k uzavření vápníkových uvolňovacích kanálů v sarkoplazmatickém retikulu. Kromě toho přepraví druhá řada dopravních pump aktivního vápníku tento vápník ze sarkoplazmy zpět do sarkoplazmatického retikula, kde kalzekvestrin, protein vážící vápník, vyluhuje vápník z roztoku a způsobí, že ještě více vápníku je vyděleno dovnitř sarkoplazmatického retikula. Tento mechanizmus je tak účinný, že koncentrace vápníku v sarkoplazmě uvolněného svalového vlákna je 10 000násobně nižší než uvnitř sarkoplazmatického retikula.
Architektura svalu
Sval je vysoce organizován nejen na mikroskopické úrovni, ale i v rovině celkové morfologie. Kosterní sval je částí složité soustavy kostí, vazů a šlach, která musí fungovat jako celek podle biomechanických zákonů (Lieber a Friden, 2000). Ačkoli jsou si sval a přídavné tkáně velmi podobné v celém živočišném světě, musí být celý systém navržen tak, aby účinně fungoval jako celek v každém specifickém druhu organizmu. Pochopení architektury svalu vyžaduje znalost strukturálních vlastností, jež ovládají funkce celých sad svalů (Lieber a Friden, 2000). Podobnosti i odlišnosti, jež existují, jsou v první řadě výsledkem biomechaniky, požadavků plánu a jeho omezení, nikoli původu ze společného předka.
Druhy svalů
U zvířat existují jen tři základní druhy tkáňových stahových soustav. V první se účastní aktin a další vlákna uvnitř buněk a způsobují jejich pohyb, třeba u prvoků. Dalším druhem jsou relativně nespecializované stahové buňky u hub a jistých složitějších zvířat (Dorit, Walker a Barnes, 1991, str. 214). Posledním druhem je svalová soustava popsaná v tomto článku. Tyto tři soustavy jsou velmi odlišné, a neexistují důkazy evoluce jedné v druhou.
A dále, neexistuje žádný empirický důkaz evoluce svalů, ačkoli bylo napsáno mnoho studií pokoušejících se ukázat homologii mezi různými formami svalů (Itina, 1979; Johnston et al., 2004; Laing et al., 1995; Meedel et al., 1997; Fukuzawa et al., 2001; McGuigan et al., 2004; Ferry-Graham a Lauder, 2001; Clayton et al., 1988; a Gibson, 1986). I když existují jisté variace, jsou všechny známé druhy svalů stejně složité jako soustava popisovaná v tomto článku.
Konkrétní zvíře vyžaduje zabudovanou flexibilitu určitých rysů (jako třeba délky svalového vlákna), aby se mohlo přizpůsobit specifickým změnám prostředí. Johnston et al. (2004) našli důkazy toho, že ačkoli je regulace produkce svalové trubice u ryb pod velkým selekčním tlakem, existuje zde optimální velikost vlákna, jež se mění přímo s velikostí těla. Tato optimální velikost je výsledkem vyrovnávání mechanických a jiných tlaků na průměr vlákna a energetických nákladů nutných k udržení požadované hladiny spádu iontů. Tento poznatek nám pomáhá pochopit, jak se život může přizpůsobovat různým prostředím, ale pouze v jistých mezích. Na rozdíl od některých názorů se zde nejedná o důkaz makroevoluce, ale o důkaz naplánované soustavy, jež umožňuje adaptaci na omezené změny prostředí.
Někteří badatelé se pokusili využít rozdílů ve svalech (jež existují z ryze funkčních důvodů) ke spekulacím o evolučním vývoji. Hoh (2002) do své studie, kde srovnává svaly obratlovců, zapracoval spekulaci o možné evoluci svalů. Otevřeně však připustil, že jde o pouhou teorii, jež není založena na přímých důkazech .
Mastropaola (2001) neshledal důkazy evoluce svalů v matematické analýze teorie vývoje svalu s maximálním hnacím podnětem, ale spíše nalezl důkazy pro plán. Konstatoval, že „lidský sval byl pečlivě do podrobností vymyšlen tvůrcem“ a že „logickým závěrem vědeckého bádání je, že sval a všechny ostatní živé tkáně byly naplánovány“ (Mastropaola, 2001, str. 219).
Svaly zachované ve fosiliích
Ačkoli měkká tkáň nebývá ve fosilních nálezech dobře zachována, svalová tkáň je zachována v některých situacích použitelně, jako třeba ve vzorcích hmyzu v jantaru. O jantaru víme, že pomáhá uchovat podrobnosti až do detailní struktury buňky, včetně struktury buněčných organel (Henwood, 1992). Měkké tkáně, „zejména letové svaly“, jsou často zachovány u zvířat pohřbených v jantaru nalezeném v Dominikánské republice (Henwood, 1992). Dominikánský jantar je většinou datován od svrchního eocénu k nejspodnějšímu miocénu.
Organela = struktura uvnitř buňky plnící specializované funkce. Jádro buňky uchovává dědičnou informaci, mitochondrie slouží zisku energie, v endoplazmatickém retikulu se na ribozómech tvoří proteiny. Na stavbě mnohých organel se podílí membrána velmi podobná buněčné membráně. Zdrojem této membrány je Golgiho komplex (pozn. překl.).
Svalová vlákna byla také nalezena u hmyzu zachyceného v jantaru baltském, o němž se soudí, že se „utvořil před zhruba 40 milióny let“ (Poinar a Hess, 1982, str. 1241). Hmyz má „složité svalstvo, a má přibližně dvakrát tolik svalů než savci“ (Grimaldi et al., 1994, str. 7). Techniky hodnocení tkáně pohřbené v jantaru jsou stále nedokonalé, ale průběžně se zlepšují. Jasně vymezené svalové pásy a další zatím prostudované struktury „připomínaly současné tkáně dehydrované etylenglykolem“, roztokem užívaným k jejich konzervování (Poinar a Hess, 1982, str. 1242).
Svalová tkáň může být také použitelně zachována u zvířat chycených v asfaltových jezerech či zamrzlých v ledu. Pár důkazů nyní existuje o uchování svalů dinosaurů. Měkká tkáň byla objevena v dinosauřích kostech, jež jsou údajně skoro 70 miliónů let staré (Wilford, 2005; Nance, 2005). Přes dvacet centimetrů dlouhé mládě teropoda nalezené ve vápencovém nánosu blízko Neapole, Itálie, obsahovalo svalovou tkáň v hrudní oblasti (Dal Sasso a Signore, 1998).
V ještě ne úplně pochopeném procesu „mohou být zachovány ve fosfátových nerostech žábry a svalová tkáň a dokonce i buněčná jádra …ryb, jež jsou 120 miliónů let staré“(Palmer, 1994, str. 17). Nejenže byly nalezeny balíky svalové tkáně, ale byla též rozlišena jednotlivá svalová vlákna. V jedné studii o nejslavnějším nálezu šlo o „měkkou, čerstvě vypadající tkáň uvnitř stehenní kosti druhu Tyranosaurus rex“ včetně krevních cév z hladkého svalstva (Yeoman, 2006; Fields, 2006). Výzkumný tým konstatoval, že „cévy i jejich obsah jsou ve všech ohledech podobné krevním cévám získaným z kostí současného pštrosa“ (Schweitzerová a tým, 2005, str. 1952).
Všechny staré svalové struktury, které byly zatím hodnoceny, jsou blízké naprosté shodě se svaly moderními. Henwood (1992) konstatoval, že malé rozdíly mezi moderním a fosilním letovým svalem hmyzu, jež byly zkoumány, jsou „nejlépe vysvětleny tafonomickými okolnostmi“ (str. 206) – jinými slovy, rozdíly jsou způsobeny změnami nastalými v procesu konzervace (jako třeba ztrátou vody), a ne evolucí svalové tkáně.
Tafonomie = obor paleontologie zabývající se vznikem, způsobem zachování a uložení zkamenělin (pozn. překl.).
Svazky „svalových vláken ve svých původních začátcích a úponech“ – a dokonce i linie Z a M a kanálky T se „zarážející věrností jako živé – byly objeveny v mnoha vzorcích dominikánského jantaru“ (Grimaldi et al., 1994, str. 1, 7-8, 10). Vzorky dosud prozkoumané ukazují, že nejstarší známé vzorky svalů ve srovnání se vzorky moderními svědčí o tom , že anatomie a fyziologie svalů se nezměnila.
Závěr
Všechny druhy svalů u všech zvířat jsou velmi podobné ve struktuře i funkci a všechny jsou nezjednodušitelně složitými stroji, které musí fungovat jako část složitého, dobře nastaveného provázaného systému, jak jsme popsali v tomto článku. Sval je součástí velmi složité soustavy, která mohla fungovat jen tehdy, byla-li přítomna každá ze základních složek ode dne jedna. Muselo jít o plně funkční soustavu od samého začátku, a bez funkčního svalu mohlo existovat jen velmi málo živočichů, od hmyzu po savce.
Toto zjištění je v příkrém rozporu s darwinovskou předpovědí, že primitivní život užíval primitivní svalstvo a pokročilejší formy života užívaly vyvinutější a složitější svalstvo. Žádné jednoduché svaly neexistují; spíše všechny svaly veškerých živočichů, od jednoduchých po složité, obsahují tytéž složky a tentýž základní plán, který jsme popsali v našem článku. Neexistují důkazy vývoje svalů ve fosilních nálezech. Svaly zůstávají stále stejné od svého počátku, v každém druhu živočicha.
Poděkování
Děkuji Jonathanu O´Quinnovi, MA DVM, Waynu Frairovi, Ph.D., Cliffordu Lillovi, MA, Georgu Howemu, Ph.D., Jodymu Allenovi, R.N., svému ilustrátorovi B.L.Lindleymu-Andersonovi a dvěma anonymním informátorům.
Odkazy na literaturu:
CRSQ: Creation Research Society Quarterly.
Bergman, J. 1999. ATP: the perfect energy currency for the cell. CRSQ 36:2–9.
Clayton, L., F.C. Reinach, G.M. Chumbley, and A.R. MacLeod. 1988. Organization of the hTMnm gene. Implications for the evolution of muscle and non-muscle tropomyosins. Journal of Molecular Biology 201:507–515.
Dal Sasso, C., and M. Signore. 1998. Exceptional soft-tissue preservation in a Theropod dinosaur from Italy. Nature 392:387.
Dorit, R.L., W.F. Walker Jr., and R.D. Barnes. 1991. Zoology. Thomson, New York, NY.
Ferry-Graham, L.A., and G.V. Lauder. 2001. Aquatic prey capture in ray-finned fishes: a century of progress and new directions. Journal of Morphology 248:99–119.
Fields, H. 2006. Dinosaur shocker. Smithsonian 37(2): 50–55.
Fukuzawa, A., J. Shimamura, S. Takemori, N. Kanzawa, M. Yamaguchi, P. Sun, K. Maruyama, and S. Kimura. 2001. Invertebrate connectin spans as much as 3.5 μm in the giant sarcomeres of crayfish claw muscle. The EMBO Journal 20:4826–4835.
Gibson, G.A.P. 1986. Evidence for monophyly and relationships of chalcidoidea mymaridae and mymarommatidae hymenoptera terebrantes. Canadian Entomologist 118:205–240.
Grimaldi, D., E. Bonwich, M. Delannoy, and S. Doberstein. 1994. Electron microscopic studies of mummified tissues in amber fossils. American Museum Novitiates 3097:1–31.
Henwood, A. 1992. Exceptional preservation of dipteran flight muscle and the taphonomy of insects in amber. Palaios—The Society for Sedimentary Geology 7:203–212.
Hoh, J.F.Y. 2002. Review: “Superfast” of masticatory myosin and the evolution of jaw-closing muscles of vertebrates. The Journal of Experimental Biology 205:2203–2210.
Itina, N.A. 1979. Evolution of muscle function: comparison of striated muscle characteristics in tunicates and vertebrates. Zhurnal Evoliutsionnoi Biokhimii I Fiziologii 15:401–413.
Johnston, I.A., M. Abercromby, V.L.A. Viera, R.J. Sigursteindottir, B.K. Kristjansson, D. Sibthorpe, and S. Skulason. 2004. Rapid evolution of muscle fibre number in post-glacial populations of arctic charr salvelinus alpinus. The Journal of Experimental Biology 207:4343–4360.
Laing, N.G., S.D. Wilton, P.A. Akkari, S.Dorosz, K.Boundy, C. Kneebone, P. Blumbergs, S. White, H. Watkins, D. Love, and E. Haan. 1995. A mutation in the α tropomyosin gene TPM3 associated with autosomal dominant nemaline myopathy. Nature Genetics 9:75–79.
Lieber, R.L., and J. Friden. 2000. Functional and clinical significance of skeletal muscle architecture. Muscle and Nerve 84:1647–1666.
Marieb, E. 2005. Anatomy and Physiology. Pearson, San Francisco, CA.
Mastropaolo, J. 2001. The maximum-power stimulus theory for muscle. CRSQ 37:213–220.
McGuigan, K., P.C. Phillips, and J.H. Postlethwait. 2004. Evolution of sarcomeric myosin heavy chain genes: evidence from fish. Molecular Biology and Evolution 21:1042–1056.
Meedel, T.H., S.C. Farmer, and J.J. Lee. 1997. The single MyoD family gene of Cona intestinalis encodes two differentially expressed proteins: implications for the evolution of chordate muscle gene regulation. Development 124:1711–1721.
Nance, A. 2005. Perhaps a step toward dinosaur DNA. New York Times National, Friday, March 25, p. A15.
OOta, S., and N. Saitou. 1999. Phylogenetic relationship of muscle tissues deduced from superimposition of gene trees. Molecular Biology and Evolution 16:856–867.
Palmer, D. 1994. How busy bacteria turn into stone. New Scientist 1917:17.
Poinar, G.O., Jr., and R. Hess. 1982. Ultrastructure of 40 million year old insect tissue. Science 215:1241–1242.
Saladin, K. 1998. Anatomy and Physiology. McGraw and Hill, New York, NY.
Schweitzer, M.H., J.L. Wittmeyer, J.R. Horner, and J.K. Toporski. 2005. Soft-tissue vessels and cellular preservation in Tyrannosaurus rex. Science 307:1952–1954.
Seeley, R., T. Stephens, and P. Tate. 2003. Anatomy and Physiology. McGraw Hill, New York, NY.
Stokstad, E. 2005. Tyrannosaurus rex soft tissue raises tantalizing prospects. Science 307:1852.
Vogel, S. 2001. A Natural History of Muscle. Norton, New York, NY.
Wilford, J.N. 2005. Dinosaur find takes scientists beyond bones. New York Times National, Friday, March 25, pp. A1, A15.
Yeoman, B. 2006. Schweitzer’s dangerous discovery. Discovery 27:37–41 and 77.