Revize knihy o Homologii, genech a evoluční inovaci od Güntera P. Wagnera
Princeton University Press, Princeton, NJ, 2014
Z creation.com přeložila Martina Potužníková, editace Pavel Kábrt – 11/2017. Translation granted by Creation.com – Přeloženo s povolením od Creation.com.

Autor: Günter P. Wagner
Tato kniha se soustředí na prolínání genetiky a embryologie; jde o tzv. vývojovou genetiku— ta se snaží pochopit, jak jsou řetězce DNA přestavěny ve fungující orgány a stavební plány těla. Naše pochopení tohoto dech beroucího komplexního fenoménu je stále extrémně kusý, ačkoliv moderní techniky konečně umožnily zahájit výzkum. Konkrétní technika mapuje, kde a kdy se v embryu projeví určitý gen.
Jiná technika potlačí (nebo ‘vyřadí’) určitý gen a poté pozoruje, které embryonální znaky se vyvinou a které ne. Většina Wagnerovy knihy (~70 %) je materiál tohoto typu a může být přijímána kýmkoli (evolucionistou nebo kreacionistou). Upřímně nevidím žádný důvod o tomto materiálu pochybovat. Nicméně jedná se o materiál velmi technický—tedy pomalé a únavné čtení;není tedy vhodný pro každého. Jako typický příklad uvedu:
“Ve fázi 40, zárodky hrudní ploutve vykazují klasické rané kolineární exprese genu HoxD. HoxD13 se exprimuje na zadním okraji zárodku ploutve, uložený mezi poněkud rozsáhlejší expresní doménou HoxD12, která je uložena v expresní doméně HoxD11, která se rozšiřuje ještě dále dopředu” (str. 350).
Existují kreacionističtí badatelé, pro které je toto důležitý materiál.
Tato kniha je evolucionistická, ale nikde se v ní neoznamuje nový důkaz proti stvoření nebo pro makroevoluci. Tato kniha nikdy nezavádí kreacionisticko-evoluční debatu. Existuje pro to nevyslovený důvod: evolucionisté jsou zmateni novými údaji a tím, jak je vysvětlit. Snaží se proto evoluční teorii revidovat, aby odpovídala těmto novým údajům. Tato kniha tento proces zahajuje.
“Evo-devo”1 je přezdívka pro tuto oblast, která se pokouší vysvětlit vývojovou genetiku prostřednictvím evoluční teorie. Wagner pro dosažení tohoto cíle navrhuje nové koncepty a terminologie jako možný základ pro další výzkum. (Tento materiál, který je pro čtenáře-laiky o něco přístupnější, je obsažen ve zbylých 30 % knihy.) Budu komentovat právě tento materiál.
Homologie a stavební plány
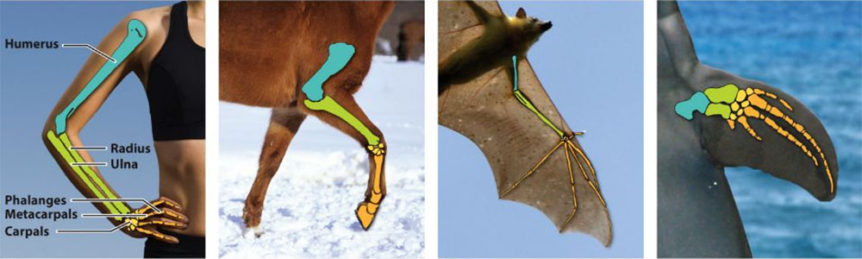
Kniha se soustředí na evolucionistický koncept homologie. Bohužel, homologie je téměř vždy definována tím, jak je problém vysvětlen, nikoli jeho vlastním pozorováním. Například Wikipedia definuje homologii jako “existenci společného předka mezi dvojicí struktur nebo genů u rozdílných druhů”. Tradičními příklady jsou ruce, tlapy, kopyta, křídla a ploutve savců, o kterých evolucionisté říkají, že jsou ‘homologické’, nebo důsledkem společného předka (viz též obrázek 1).
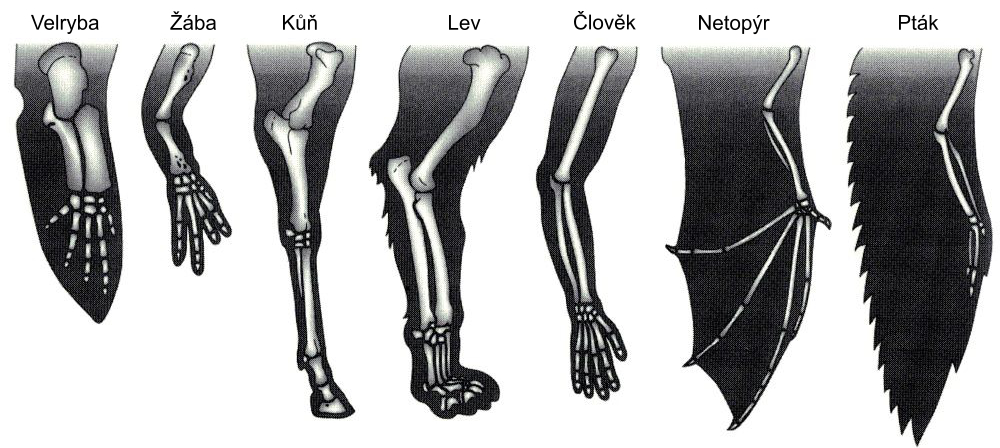
Obrázek 1. Wagnerův učebnicový příklad homologie (str. 24)
Kniha váže tři pojmy dohromady: (1) existuje sada genů; (2) ty jsou potřebné pro vypracování inovativních stavebních plánů; a (3) evolucionisté věří, že tyto geny jsou zděděny od nějakého společného předka (tj. jsou homologické). Takže Homologie, geny a evoluční inovace je vhodný název pro tuto knihu. Evoluční původ neobvyklých stavebních plánů byl vždy kontroverzní, zvláště pak v dnešní době díky vynikající knize Stephena Meyera, Darwin’s Doubt (2013) [Darwinova pochybnost],),2 o náhlém výskytu různorodých stavebních plánů během kambrické exploze fosilních živočišných forem. Wagner připouští, že původ nových vlastností a nových stavebních plánů “je jednou z nejdůležitějších a nejméně prozkoumaných otázek v evoluční biologii” (str. 3). Fosilní záznamy a kambrická exploze ukazují pouze podobu a tvar orgánů a stavebních plánů (tzv. morfologii). Wagner uznává, že se jedná o “jednu z nejsložitějších tříd homologických vztahů k vysvětlení” (str. 1).
Přestože je homologie považována za hlavní důkaz evoluce, samotný koncept je problematický. Wagner poznamenává: “Neexistuje žádný konsensus, dokonce ani úzký konsensus na téma homologie a její mechanistické základy. … Každý biolog bude souhlasit, že homologie je zmatené a matoucí téma” (str. xii). “Při vysvětlování evoluce stavebních plánů nemůže být dosaženo žádného trvalého pokroku bez důkladné reorganizace. Problémem je, že mnoho protichůdných stanovisek k homologii dávalo smysl v rámci výzkumných programů, ve kterých byly představeny [zdůraznění dodáno]” (str. 5). Takže Wagner usiluje o aktualizaci konceptu homologie tak, aby byl vhodný zvláště pro evo-devo výzkum.
Hlavní problémy
Kniha naznačuje rozličné problémy čelící evo-devo. Například:
- “V současné době je dobře zavedené a běžné, že všechna zvířata sdílejí soubor konzervovaných genů, které jsou přirozeně důležité pro vývoj charakteristiky stavebních plánů těla. Jednalo se o hluboce překvapivý objev, protože v tradici neo-darwinistické evoluční biologie byla možnost homologických genů mezi vzdáleně příbuznými druhy výslovně vyloučena [zvýraznění přidáno]” (str. 26).
Wagner odvádí špatnou práci při odhalování tak podstatného problému. Proto ho vysvětlím zde. Mnoho z důležitých genů stavebních plánů těla je široce sdíleno mezi různorodými živočišnými kmeny. Pro evolucionisty to znamená, že tyto geny musely existovat uvnitř nějakého starodávného společného předka všech těchto skupin. Ale tyto stavební plány jsou docela odlišné jeden od druhého, tak odlišné jako obratlovci, hvězdice, medúzy a hmyz. Proto musel tento společný předek existovat mnohem dříve než první výskyt těchto skupin—mnohem dříve než kambrická exploze. Jinými slovy, tyto velmi důležité geny stavebních plánů musely vzniknout v době, kdy existovaly mikroorganismy a relativně málo co s tělem. Jak by mohly geny, vzniklé v mikroorganismech nebo třeba v původních dávných medúzách být dnes podstatným základem pro současné, zcela nesourodé stavební plány v různých tělech? Je zcela hloupé tvrdit, že přirozený výběr tyto geny vytvořil původně k ovládání rozličných tělních plánů: (1) protože onen dávný předek (ať už to bylo cokoliv) neměl mnoho z toho, co nazýváme tělo; (2) protože přírodní výběr nemůže vytvořit něco pro budoucí použití; a (3) je nepravděpodobné, aby se tyto geny při svém vzniku jen tak vytvořily, prostě náhodně tak, že by se pak hodily v budoucnu k využití pro stavební plány zcela odlišných těl.
Tento problém způsobuje revoluci v evolucionistickém uvažování. V současnosti mnoho evolučních specialistů prohlašuje, že přirozený výběr je nedostatečný, a je třeba nějakých dodatečných vysvětlení.3,4
I Wagner se podobně domnívá, že přírodní výběr je nedostatečným vysvětlením. 5 Jako dodatečné vysvětlení pokládá důraz na strukturalismus.6 Strukturalismus se zaměřuje na formu (nebo strukturu) a říká, že toto klade omezení na typy změn, ke kterým může dojít. A tak je evoluce omezena strukturou organismu a to omezuje směr, kterým se evoluce může vydat. Některé znaky jsou zachovány ne pro jejich funkčnost, ale proto, že jsou vestavěny do struktury organismu. V jistém smyslu je evoluce s nimi pevně ‘svázána’.
To je ale v rozporu se selekcionistickým vysvětlením, které argumentuje tím, že daný znak byl zachován nebo ztracen přesně kvůli svému efektu na funkčnost. Také je to v konfliktu s vysvětlením neutralistů, které tvrdí, že daný znak byl zachován anebo ztracen právě proto, že neměl žádný účinek na funkci. Všechny typy vysvětlení–strukturalistické, selekcionistické, neutralistické a mnohé další—jsou součástí evolucionistova rozsáhlého, chaotického a nejrůznějšími teoriemi naplněného “švédského stolu spekulací”—kde je každý nápad vybrán či zavržen dle momentální evolucionistovy potřeby.
No a druhý problém překvapil evolucionisty: homologické znaky nejsou často způsobeny homologickými geny.
- “Nicméně problematický je fakt, že homologické znaky mohou zjevně pocházet z odlišných vývojových mechanismů u rozdílných druhů” (str. 37). “Vzrůstají důkazy toho, že homologické znaky ze vzdáleně příbuzných organismů, jako kobylky a octomilky, často používají dosti odlišné geny pro vývoj jednoznačně homologických znaků, jako jsou články hmyzího těla. A proto nemohou být identické morfologické znaky vysvětleny identickou sadou genů, které vedly k jejich vývoji [zvýraznění dodáno]” (str. 2).
- “… nejobtížnější problém, když se pokoušíme vysvětlit shodnost znaku; totiž nepopiratelné homologie (čili identické znaky napříč druhy) je často spojen s rozsáhlými variacemi při vývoji biochemických cest a mechanizmů, které tyto znaky produkují [zvýraznění dodáno]” (str. 6). “Existuje rostoucí počet důkazů … které ukazují, že zdánlivě ten stejný znak může být realizován pomocí rozdílných genů v rozdílných druzích” (str. 74).
- “Kontinuita morfologických znaků není podporována kontinuitou genetické informace. To vypadá na docela pěkně depresivní situaci … a motivuje myšlenku, že homologie by mohla být iluzí … [zvýraznění dodáno]” (str. 90).
- “… vývojové biochemické cesty homologických znaků se mohou podstatně měnit mezi různými druhy bez toho, že by to ovlivnilo identitu daných znaků.” (str. 412).
- “… podobnost genové regulační sítě některých tkání, dokonce takových, kde je samo o sobě zahrnuto mnoho genů, není dostatečně silným důkazem homologie ” (str. 113).

Obrázek 2.
Obrázek 2. Fosilie želvy z Wagnerovy knihy. Wagner píše: “Toto je problém darwinistického způsobu myšlení: jaká evoluční sekvence může vysvětlit původ plánu tělesné stavby želvy v řadě malých kroků, když lopatka může být pouze vně nebo uvnitř hrudního koše? Jaké by byly mezistupně? Záležitosti nepomůže ani fakt, že fosilní záznamy neodhalují žádné přechodné morfologie. Želvy se zdají být definitivně nadějnými monstry—náhlá, radikální odchylka od plánu tělesné stavby předchůdce bez jakýchkoli přijatelných nebo dokumentovaných přechodných forem. Co dále ztěžuje situaci je to, že je dokonce obtížné přiřadit jejich fylogenetickou příslušnost k fosilním formám” (str. 180).
Zkrátka mezi morfologií a jejími výchozími geny existuje značná nezávislost nebo nesouvislost—nejsou navzájem homologické. Tím je homologický koncept narušen. Co je tedy skutečná homologie: je to morfologie nebo geny, co určuje homologii?
A je to ještě horší. Jak se embryo vyvíjí, objevují se elementárně odlišné typy buněk, množí se a nakonec formují rozličné tělesné orgány. Evolucionisté tradičně chápali tyto rozličné typy buněk7 jako důležitý klíč k homologii orgánů, které produkují. Wagner naznačuje, že tento pohled byl vyvrácen moderními důkazy. Wagner píše, že existuje,
- “… řada příkladů, u nichž jasně homologické znaky vznikly buď v průběhu embryogeneze z různých populací buněk, nebo sledovaly různé vývojové cesty, aby pak dospěly k té samé formě mofrologie v dospělosti jedince. Jedna z možných reakcí na tento fakt je tvrzení, že celý koncept homologie je nesmysl. [zdůraznění dodáno]” (str. 90).
Nicméně Wagner se drží “druhé možnosti, a sice předpokladu, že třeba embryologické počátky jsou pro základy homologického vývoje nepodstatné … [zdůraznění dodáno]” (str. 90).
Takže pro Wagnera geny nejsou spolehlivé a embryologické počátky nejsou relevantní jako indikátory homologie. Představuje proto nový koncept zahrnující ‘sítě’ genetického materiálu. Jeho koncept nazývaný Character Identity Network nebo ChIN,8 [síť identity znaků], slouží jako možný zdroj homologie, protože nic z ostatních věcí není spolehlivým zdrojem. Tento koncept je dosud mlhavý a nejasný. Zdá se, že jeho účelem je povzbudit ‘výzkum’ a podnítit spřádání nových vysvětlování.
Homologie versus konvergence
Homologie byla dlouho považována za hlavní důkaz ve prospěch evoluce. Bohužel, evolucionisté ji obvykle definují na základě vysvětlení, nikoli na základě pozorování. To vyvolává otázku: jak můžeme homologii pozorovat? Bez jasného způsobu, jak to udělat, jsou evolucionisté nebezpečně blízko k prohlášení: “Společné podobnosti mezi rukama, tlapami, kopyty, křídly a ploutvemi jsou homologie, a to proto, že my evolucionisté říkáme, že jsou homologiemi—a homologie je z definice dána společným předkem.
A tak evoluce je fakt! ”Potřebujeme způsob, jak protnout takový argument odvážným výrokem nebo pomocí definice kruhem. Zejména, jak pozorujeme nebo identifikujeme homologii? Může to být provedeno objektivně, bez toho aniž bychom se opírali o to, co říkají evolucionisté?
Zde je hlubší problém. Když dva druhy sdílejí podobný znak, pak by to možná mohlo být vysvětleno společným původem (tj. homologií), ale často to společným původem vysvětlit nelze. Evolucionisté tyto druhotné znaky nazývají ‘konvergence’ nebo ‘homoplazie’, a to je v přírodě velmi časté—na morfologické, embryologické a molekulární úrovni.9 Příkladem je vaše oko a oko chobotnice—které jsou pozoruhodně podobné –obě obsahují čočku, duhovku, sítnici, optický nerv a svaly pro rotaci oka a ovládání duhovky—avšak tyto podobnosti nelze vysvětlit společným původem.
Evolucionisté rozdělují podobné znaky do dvou typů: ty, které mohou být vysvětleny společným původem a ty, které ne. Evolucionisté zpravidla mlčky odsouvají druhý typ, vypouštějí ho z diskuzí, jako kdyby byl irelevantní. To dělá i Wagner. Pokouší se objasnit koncept homologie jako ústřední námět své knihy, přesto sotva zmiňuje konvergenci. Toto opominutí vyvrací jakýkoli pokus o vysvětlení.
Prezentace zaměřené na přesvědčení široké veřejnosti, že evoluce je ‘fakt’ drasticky zlehčují konvergenci nebo ji zcela opomíjejí. Tento přístup—zametání konvergence pod koberec— je evolucionisty využíván, když se vás snaží přesvědčit, že ‘v evoluční teorii je všechno v pořádku’. Wagner se v tomto režimu nachází, když brouzdá novými daty z vývojové genetiky—které evolucionisty matou. Téma konvergence by pro Wagnerovy účely bylo nepříjemností. Také by pokazilo jeho pokus o objasnění konceptu homologie. Takže se mu tiše vyhnul.
Všechny důkazy podporují evoluci?
Porovnejte to s knihou z roku 2004, Life’s Solution: Inevitable Humans in a Lonely Universe [Řešení života: Nevyhnutelní lidé v osamělém vesmíru], kde Simon Conway Morris poprvé bere evoluci jako nepopiratelný fakt, a poté se zaměřuje výhradně na konvergenci, a cituje mnoho pozoruhodných příkladů. Například, podle evolucionistů se počátek zraku objevil ve více než 40 odlišných dobách (jako složené oko mouchy) a počátek oka s čočkou nejméně v sedmi různých obdobích–u obratlovců, hlavonožců (tj. chobotnic), medúz, pavouka, kroužkovců a korýšů. Morris poté používá konvergenci (a její hojnost) jako důkaz síly přírodního výběru. Proto dochází k závěru: pokud život existuje jinde ve vesmíru, potom bude (díky konvergenci) téměř jistě podobný životu na Zemi a pravděpodobně bude zahrnovat bilaterální vzpřímené hominidy s velkými mozky, jako jsme my. Jeho kniha je plná důkazů svědčících proti evoluci, přesto evoluci považuje za nepopiratelný fakt.10
Zde je důvod. V evolucionistickém způsobu myšlení nemůže existovat žádný důkaz proti evoluci. Místo toho jsou všechny důkazy proti evoluci odvysvětleny jako důkazy ve prospěch některých z evolučních vysvětlení—nehledě na to, jak jsou přitažené za vlasy nebo na nedostatek experimentálních důkazů nebo na to, jak jsou netestovatelné—jednoduše proto, že evoluce je ‘fakt’! Například existují podobné znaky, které nemohou být vysvětleny společným původem,11 a ty jsou v přírodě velmi hojné—což je samo důkazem proti evoluci. Místo toho evolucionisté vše interpretují jako důkaz ve prospěch ‘neuvěřitelné síly přírodního výběru’.
Stejně chybné myšlení se vyskytuje mnohokrát: u velkých morfologických mezer ve fosilních záznamech (tj. nedostatek pozvolných kroků ve velkých měřítcích); u systematické absence jednoznačných předchůdců a rodokmenů; u bičíků bakterií; u biomolekulárního modelu nazývaného ‘synchronizovaná evoluce’; a u původu života, abychom jmenovali alespoň něco. Tento typ myšlení zkresluje evolucionistické formulace kdekoliv.
Syrová, spekulativní evolucionistická vysvětlení jsou podávána jako ‘hodnověrná’, ‘důvěryhodná’, ‘uspokojivá’, nebo prostě jako ‘fakt’. Prakticky všechny evolucionistické knihy, včetně Wagnerovy, zapadají do této šablony myšlení. To se odráží v evolucionisty často opakovaném sloganu: ‘Evoluce je fakt; akorát si, my evolucionisté, nejsme jisti, jak k ní vlastně došlo.’
Objektivní homologie
Evolucionisté potřebují nějakou objektivní metodu pozorování a rozlišování homologie od konvergence. Často užívanou metodou je použití kladistiky. Kladistika zařazuje druhy do skupin hierarchicky uspořádaného vzoru, který se jmenuje kladogram. Je to jako klasifikace knih v knihovně, kde jsou knihy vědecké, z nich některé jsou o fyzice, z nich některé jsou o nukleární fyzice a tak dále v soustavě podmnožin a jejich podmnožin a jejich podmnožin—jako matrjošky. Kladistika zejména, usiluje o vytvoření klasifikace, která maximalizuje počet vnořených znaků a minimalizuje počet nevnořených (obsažených vs. neobsažených) znaků. Tato metoda (známá jako transformovaná kladistika) je vědecky neutrální (a použitelná jak kreacionisty, tak evolucionisty) a neumožňuje evolucionistickým fantaziím deformovat výsledky. Závěry jsou vysvětleny později.
Poté, co je identifikován nejlepší kladogram, evolucionisté vysvětlují nezařazené znaky jako ‘konvergenci’ a vnořené znaky jako ‘homologie’. Kladistika zkrátka automaticky minimalizuje počet ‘konvergencí’ a maximalizuje počet ‘homologií’. Jenže transformovaná kladistika moc evolucionistům nepomáhá. Proto vytvořili verzi zvanou evoluční kladistika (nebo klamně nazývaná ‘fylogenetická kladistika’12), která umožňuje evolucionistickým báchorkám modifikovat výsledek. Evolucionisté tak mohou změnit výsledek pomocí vysvětlení ztrát, kde se tvrdí, že daný znak byl v některých organismech ztracen, zatímco v jiných zachován. Někteří evolucionisté (včetně Wagnera) také tvrdí, že znak se může znovu vyvinout poté, co byl ztracen. Tyto spekulativní scénáře mohou změnit topologii kladogramu, čímž se změní identifikace konvergencí a homologií.13
Jinými slovy, používání evolucionistických báchorek dává evolucionistům určitou volnost reklasifikovat konvergence a homologie různými způsoby—v závislosti na jejich potřebách v konkrétní chvíli. Mají určitou flexibilitu.
Přesto pro Wagnerovy účely není tato flexibilita dostatečná, a proto odmítá jakékoli striktní spoléhání na kladistiku a považuje ji za nekompatibilní s jeho výzkumným programem.14 Nezbývají mu tedy žádné nezávislé nástroje k identifikaci homologie. Namísto toho jsou kladistické objevy považovány za zcela sugestivní: jsou v závislosti na potřebách evo-devo báchorek přijímány nebo odmítnuty.
Wagnerovým řešením je definovat homologii neurčitě, což ve skutečnosti umožňuje evo-devo badatelům ji znovu a znovu definovat od případu k případu. Wagner říká: “[Budeme] se snažit doplnit biologické detaily a co tím myslíme, když říkáme, že jsou dva znaky stejné [zdůraznění je v originálu]” (str. 244–245). Jinými slovy, dva znaky by mohly být ‘stejné’ (a tedy homologické) na základě morfologie, nebo genů, nebo typu embryologických buněk, nebo ChIN, nebo na základě čehokoli jiného, co se badatel rozhodne chápat jako ‘biologický detail’. Je to široce otevřený koncept. Wagner trvá na tom, že: “upustit od hledání definice homologie nemusí být ke škodě [zdůraznění je v originálu]” (str. 244).
Wagner chce v evo-devo výzkumu pokračovat. Proto se domnívá, že je lepší mít neurčitě definované pojmy než žádné pojmy.
Problém oddělený versus sdílený
Na další problémy narazíme při pokusu evolucionistů vysvětlit specifické stavební plány těl. Zde jsou některé běžné příklady: (1) Přírodní výběr nemůže znak zdokonalit, pokud jeho genetika není vystavena výběru—nebo ‘individualizovaná’15— a to vyžaduje, aby genetika daného znaku byla oddělená a odlišná od ostatních věcí. Na druhou stranu, (2) evolucionisté často chtějí, aby zdokonalení byla sdílena na mnoha místech těla—takže zdokonalení designu (např. ploutve, peří, končetiny, šlachy nebo oka) by se objevilo na všech takových pozicích daného těla—a to by vyžadovalo nějaký prostředek sdílení stejných genetických instrukcí mezi všemi (a pouze těmito) pozicemi daného designu. Ale to jsou dva protichůdné požadavky—oddělené genetické instrukce versus sdílené genetické instrukce. Nemůžete mít oba současně. Proto evolucionisté potřebují nějaký prostředek pro přepínání tam a zpět podle toho, jak zrovna jejich báchorka vyžaduje. Rovněž genetická reorganizace (jak při sdílených instrukcích, tak při oddělených) musí být přesně cílena aby se vyhnula narušení zbytku genomu.
Předpokládejme například, že chcete vysvětlit evoluční původ rybích ploutví. Můžete si představovat, řekněme, že přírodní výběr nejprve vytvořil genetické instrukce pro primitivní ploutev, následované pak instrukcí “udělej to čtyřikrát” (s bilaterální symmetrií), aby vytvořil dvě přední primitivní ploutve a dvě zadní primitivní ploutve. V tomto okamžiku není možné specializovat některé ploutve samostatně bez ostatních, protože “udělej to čtyřikrát” nevystavuje jednotlivé ploutve výběru. Aby byla specializace předních ploutví odlišná od zadních ploutví, musí být nejprve genetika ploutví reorganizována tak, aby umožňovala samostatnou genetiku—aby mohly projít samostatně výběrem. Sama o sobě není taková genetická reorganizace adaptivní, protože nemá okamžitou výhodu. A vzhledem k tomu, že přirozený výběr nemůže vybírat pro budoucí použití, musí se daná genetická reorganizace objevit bez významného vedení přírodním výběrem. Navíc jakmile k tomu dojde, nelze vysvětlit sdílené změny všech čtyřech ploutví, protože nyní mají samostatnou genetiku.
Na podobný problém narazíme v případě peří, kde je sdílený plán pera použit po celém těle, přesto je každé pero specializováno pro nějaký účel (např. letky pro létání versus prachové peří pro zahřátí), délku a barvu.
Tímto způsobem může jeden evolucionistický výklad proměnit další evolucionistický výklad ve výklad neobratný nebo nepravděpodobný. Nakonec tak evolucionistické výklady zakopávají o vlastní nohy a zamotají se do vlastních protichůdných požadavků. Tento typ problému se v evo-devo vyskytuje často.
Evo-devo výklad potřebuje přesně cílenou genetickou reorganizaci, tam a zpět mezi oddělenými genetikami versus sdílenými genetikami. Vznik pečlivě sestrojené rybí ploutve—plus jejich pozdější přeměna v ruce, tlapy, kopyta, křídla a ploutve savců—by to vyžadoval. Vaše ruce jsou vysoce specializované zcela odděleně od vašich chodidel, přesto vaše ruce a chodidla sdílejí fakt, že mají pět prstů—zcela odlišně od rybích ploutví. Podobně je to i u křídel a nohou netopýrů. Podobně u ploutví velryb. A tak dále. Bylo by potřeba mnoho a mnoho cílených genetických reorganizací správného druhu,16 pravidelně prokládaných přirozeným výběrem správného druhu.17 Máme věřit, že příroda skáče podle toho, jak evo-devo báchorkáři pískají?
Vzájemná shoda
Aby vysvětlili historii znaku, předpokládají běžně evolucionisté, že je řízen čistě jedním genem—tedy že gen má pouze jeden účinek a žádné vedlejší efekty. A proto se jakýkoli selektivní účinek na znak (ať už je prospěšný či naopak) bude stejně vztahovat také ke genu. Na základě tohoto předpokladu jsou osud znaku a osud genu podstatně svázány jeden ke druhému.18 Tím se evolucionistické báchorky zdají mnohem více přijatelné a tak to bylo po desetiletí používáno v učebnicích evoluční genetiky, v počítačových simulacích a obzvláště v prezentacích mířených na neobeznámenou širokou veřejnost.
Tento předpoklad není přijatelný. Nyní je známo, že většina genů má vliv na více než jeden znak—tomu se říká pleiotropie. Přemýšlejme o tom jako o vedlejších efektech. Studie nyní ukazují, že průměrně každý gen ovlivňuje 7+ více různých znaků, přičemž některé geny ovlivňují až 35 různých znaků. Jak může dojít k makroevoluci, když jsou vedlejší efekty genu vzájemně provázány tolika různými způsoby? To činí makroevoluci mnohem hůře ospravedlnitelnou, a proto se obhájci evoluce tomuto problému vyhýbají a mají tendenci ho zamlčovat.19
Podobné konceptuální vyhýbání se tomuto problému se objevuje ve Wagnerově knize, kde tento druh problémů opomíjí. Používá mnoho různých pojmů pro speciální seskupení genů,20 a tak je tu jednoduše nazývám ‘genové sítě’. V klíčových bodech jeho výkladu potřebuje, aby jeho genové sítě měly pouze jeden účinek na vývoj21 —aby na vývoj neměly rozmanité vedlejší (nežádoucí) účinky—a proto jsou jeho genové sítě čistě vystaveny přírodnímu výběru (přinejmenším když chce, aby byly). Wagner opomíjí možnost, že jeho genové sítě mají rozmanité nežádoucí vedlejší účinky, které jsou mimo kontrolu evolucionistických výrobců báchorek. Jeho tichého a bezpodmínečného předpokladu je dosaženo opomíjením jakékoli vážné diskuze o problému.
Nyní prokombinujme tyto body dohromady: (1) vývoj je řízen genovými sítěmi; (2) v nich má každý gen (a navíc ještě ‘vznikající vlastnosti’ vyvíjející se genové sítě) naprosto typicky mnoho vedlejších účinků; a (3) evoluční báchorky vyžadují časté genetické reorganizace—což musí náhle vystavit tyto různé vedlejší účinky různým genetickým kontextům. Všimněte si, že tyto tři body jsou navzájem náhodné.22 Může to být něco jiného než pouhé zbožné přání, mohlo by to být něco založeného na vědeckých základech, čemu bychom mohli věřit, aby tyto procesy nebyly naprosto a pouze jen zcela zničujícící, škodlivé?
Pokud jde o stvoření
I když to Wagner nezamýšlel, jeho materiál je mimořádně povzbuzující jak pro kreacionisty, tak pro teorii informace obecně. Budu o tom diskutovat dále.
Teorie informace tvrdí, že životní formy byly rozumně navrženy, aby dosáhly současně tří cílů. Biologicky jsou navrženy: (1) pro přežití; (2) aby vypadaly jako produkt jednoho návrháře (spíše než produkt několika návrhářů působících nezávisle); a také (3) aby odolaly makro-evolucionistickým výkladům (a to všem, nejen Darwinovu). Wagnerův materiál potvrzuje bod 2 a 3 (a nepopírá bod 1).
Třeba existence sdílených genů—základních pro plány těl různých zvířat—pomáhá sjednotit životní formy jako produkt jednoho návrháře, zatímco tyto stejné geny jsou radikálním problémem pro makroevoluci. Všech tří cílů (1, 2 a 3) je dosaženo současně. To odpovídá teorii informace.
Anebo vezměte jiné údaje: homologické struktury nejsou často vyvolány homologickými geny nebo homologickou embryologií. Podobné struktury sdílené mezi druhy ukazují na to, že měly stejného projektanta. Ale často jsou způsobeny rozdílnými geny a různými embryologickými cestami, které vzdorují makroevolucionistickým výkladům. Všech tří cílů (1, 2 a 3) je dosaženo současně. To zcela odpovídá teorii informace.

Obrázek 3.
Obrázek 3. Obecný způsob, jak omezit biologickou změnu, je použít stejnou sekvenci DNA—stejný gen—různými způsoby (od Williamse 27).
Tento vzorek dat ukazuje, že evo-devo vysvětlování jsou jen bezobsažné blafy. Evo-devo výklad homologie je odhalen jako nesoudržné, beztvaré vypravěčství báchorek—kde základem pro identifikaci a vysvětlení ‘homologie’ jsou zcela rozporné případy, odporující jeden druhému. Zcela to odporuje cíli projektu 3: život byl vyprojektován tak (ve svých konkrétních částech), aby vzdoroval makroevolučním vysvětlováním.
A nakonec si vezměte Wagnerovo opětné zdůrazňování strukturalistického vysvětlování — myšlenka, že forma a struktura organismu omezuje (nebo limituje) jeho evoluci. Přesto, že to Wagner nezamýšlel, přiblížil se kreacionistickému postoji. Jako důsledek cíle číslo 3, teorie informace předpovídá, že23 životní formy byly vyprojektovány s omezeními vůči biologickým změnám.24 Jinými slovy teorie informace předpovídá strukturalismus, a to opravdu hodně strukturalismu!25
A obecným způsobem, jak omezit biologickou změnu, je použít stejnou sekvenci DNA—ten samý gen—u mnoha rozdílných biochemických cest. Tento složitý ‘matrix’ nebo mozaika propojených genů znamená, že by velké změny byly eliminovány kvůli jejich nepříznivým účinkům na přežití. Jeden příklad jsou geny, které mají četné vedlejší účinky (tj. pleiotropie, diskutovaná výše). Další příklad jsou četné geny překrývající jeden druhý (mnoha různými způsoby) na tom samém vláknu nebo na opačných vláknech dvoušroubovice DNA—tyto jevy jsou již běžně známé (Obrázek 3). Dalším příkladem je, že “existuje stále více důkazů, že stav genetické regulační sítě buňky je ovládán ne jednou centrální sítí, ale mozaikou hustě propojených síťových modulů [zvýraznění dodáno]” (str. 423). Dalším příkladem předpovězeným teorií informace je, že mnoho klíčových genů je použito stejně tak pro tělo a spermii, a stejně tak je mnoho klíčových genů použito jak pro tělo tak i pro vajíčko.26 Takže opět, ta samá DNA je použita nejrůznějšími způsoby. Struktury jako tyto pomáhají omezit změny těchto genů. Očekává se, že se objeví ještě další strukturalismus.
Kreacionisté by se neměli obávat materiálu ve Wagnerově knize. Naopak, mělo by je to povzbudit.
Odkazy a poznámky
- Rovněž se používá opačné pořadí slov, ‘devo-evo’.
- See review: Woodmorappe, J., Darwin’s dirty fossil secret, J. Creation 28(1):45–49, 2014.
- See Mazur, S., The Altenberg 16: An Exposé of the Evolution Industry, North Atlantic Books, Berkeley, CA, 2010.
- ReMine, W.R., Desperate attempts to discover ‘the elusive process of evolution’ (review of The Altenberg 16), J. Creation 26(1):24–30, 2012; creation.com/review-altenberg-16.
- Wagner je docela mlhavý (téměř to neuvádí) v otázce popisu tohoto problému a odmítá čistě selekcionistické vysvětlení. Málo kdy o těchto věcech diskutuje. Což je tajemné, protože je to ústřední hnací silou jeho knihy, proč by tedy právě zde měl mlžit? Důvodem je možná právě onen evolucionistický zvyk vyhýbat se výrokům, které by byly vodou na mlýn kreacionistům.
- Wagner také oživuje koncept typologie, který byl oprávněně zamítnut během 20. století, především Ernstem Mayrem.
- I na buněčné úrovni je homologie buněk ‘ztlumena’ (nebo rozpojena) od buněčné funkce a buněčného fenotypu (str. 270).
- Wagner ponechává, aby byl ChINs maskován a později zase odkryt, aby tak byl umožněn genetický účinek z minulosti či ‘atavismus’ (str. 418).
- Hojnost stavů nazývaných ‘konvergence’ je předpovězena informační teorií, protože to přesně odpovídá třem projekčním cílům v teorii informace.
- Viz studie: ReMine, W.R., Evidence for Message Theory, J. Creation 20(2):29–35, 2006.
- Ještě přesněji, existují sdílené podobnosti, které nelze vysvětlit společným předkem, ani atavismem, ani postranním přenosem DNA—a tak to evolucionisté nazývají ‘konvergencemi’.
- Kladisti nikdy neurčují skutečné předky. (Tady slovo ‘skutečný’ znamená, že jejich fosilie můžete držet ve svých rukách.)
- Wagner píše: “určité konceptualizace homologie, například ty, které dělá tradiční kladistická taxonomie, jsou s tímto programem nekompatibilní” (p. 4).
- Podobné vyprávění báchorek se používá vždy a často, když hrozí, že by byla podkopána objektivita evoluční kladistiky. Z toho důvodu se skupina evolucionistů včetně Colina Pattersona odtrhla, aby našla více objektivní transformovanou kladistiku.
- Wagner rezignoval na seriózní vysvětlení problému oddělenosti kontra sdílení. Velmi důkladně se tomu vyhnul. Místo toho často mluví o “individualizaci”, což ale neřeší tento problém a zatemňuje to složitost mechanizmu (například cílenou genetickou reorganizaci) nutnou k jejímu dosažení. On považuje individualizaci za tak nedůležitou, že ji dokonce vynechává i ze svého rozsáhlého indexu.
- Wagner také píše: “Novinky zřejmě vyžadují reorganizace genové regulační sítě ve velkém rozsahu” (str. 125).
- Wagner píše: “Až dosud [evoluční] výzkum původu párových končetin u obratlovců neposkytl dostatečné vysvětlení” (str. 333); a “a stále zápasíme s těmi nejzákladnějšími otázkami kolem přechodu od ploutve ke končetině” (p. 334).
- Tento evolucionista si přeje—nějak elegantně zakrýt nebo minimalizovat genetické vedlejší účinky—proto evolucionisté běžně předpokládají ‘hierarchickou homologii’, kdy genové sítě hierarchicky omezují vedlejší účinky. Wagner říká, že by se měli evolucionisté ve svém myšlení “osvobodit” od této myšlenky, protože fakta ukazují, že “homologie není hierarchická”, ale naopak “příčná” (str. 420–422). Tato ‘příčnost’ genetického vzoru— nebo mnohočetnost vztahů mezi znaky a geny—je očekávána teorií informace (viz níže). Návrat k textu.
- Na druhé straně evolucionisté aktivně lnou k pleiotropii u těch zřídkavých případů, kdy to napomáhá jejich báchorkaření.
- Wagner používá různá jména pro soubory genetického materiálu, které jsou větší než gen, a které řídí vývoj orgánů a tělních plánů, a které mohou zahrnovat pozitivní a/nebo negativní zpětnou vazbu. Používá termíny jako: genové regulační sítě, jaderný regulační komplex a identická síť znaků.
- Anebo možná dobře ohraničená skupina účinků, které zrovna náhodou souvisejí se znaky, jsou dle Wagnera vystaveny selekci.
- Také si připomeňme, že v genetice při procesu separace,—i když jde o přesné zacílení—nedochází k žádnému okamžitému bezprostřednímu selektivnímu užitku.
- Kreační teorie, jak je tradičně praktikována, vlastně nepředpovídá limity biologických změn. Místo toho jsou limity biologických změn pozorovány a tato pozorování je použito jako důkaz proti evoluci a pro stvoření.
- Přesněji, výsledek designu číslo 3 předvídá, že změny v organizmech, které lze experimentálně demonstrovat (třeba v boxech pro křížení) budou vždy mnohem menší než velké mezery v záznamu života. Tím pádem cílený plán č. 1—že životní formy jsou rozumně navrženy pro přežití—umožňuje určité schopnosti životním formám se adaptovat pro přežití v měnícím se prostředí. Teorie informace tvrdí, že je nutno vytvořit kompromis, aby šlo dosáhnout souběžně všech tří položek navrhovaného designu—a toho bylo rozumně dosaženo.
- Teorie informace předpokládá, že strukturální změny u tělních plánů (čili vlastnosti kmenů) budou mít sklon k největším omezením (obtížné dosáhnout experimentálně), a už pak volnější omezení u méně obecných plánů (tj. u vlastností tříd), a tak dále (tedy směrem k řádům a čeledím atd.). Toto pomáhá vzdorovat životu makroevolučním vysvětlováním. Nejsnáze demonstrovatelné pak budou vlastnosti druhů a rodů, čili na této úrovni, jako je barevnost nebo délka kostí končetin.
- Nejsem si vědom, že by toto bylo objeveno.
- Williams, A., Astonishing DNA complexity demolishes neo-Darwinism, J. Creation 21(3): 111–117, 2007.
Související články
- Homology made simple
- Does homology provide evidence of evolutionary naturalism?
- A serious problem for homology
- Problems with the evolutionary interpretation of limb design
Další čtení