
Část I.: Rod Homo
(Z creation.com/ přeložil M. T. – 02/2007. Druhou část naleznete zde – Část II. Hominidi nepatřící do rodu Homo.
Tento přehled analyzuje fosilní důkazy pro údajné opolidi řazené do rodu Homo. Vědci věří, že rozdíly v morfologických rysech fosilních druhů zahrnutých do rodu Homo (kromě neplatného taxonu Homo habilis) reprezentují (mezi jinými faktory) genetické variace v rámci jediného lidského rodu. O druhu Homo habilis vědci usuzují, že představuje sbírku druhově různých fosilií, jež byly buď lidské (např. Homo erectus) či šlo o australopitekní lidoopy. Pokud by fosilie jako ony řazené do Homo erectus a neandertálců byly všechny plně lidské, pak důkazy pro evoluci člověka v zásadě selhávají, jelikož existuje nepřeklenutelná morfologická mezera mezi australopitekními lidoopy a těmito lidmi.
V západní společnosti se učí a prosazuje názor (ve vzdělávání a v médiích), že člověk není přinejlepším nic jiného než vysoce vyvinutá opice. Jako trumf v tomto přesvědčování se předvádí souvislá řada údajných fosilií opolidí – poslední úder tomu, kdo by se odvážil pochybovat o této pohádce. Existují skutečně přesvědčivé fosilní důkazy o původu člověka z opice či jde prostě o další příklad jednostranné indoktrinace scientismem, materialistickou filozofií, jež požaduje přírodní vysvětlení pro všechny fenomény v kosmu (1)? Jinými slovy: je možné, že scientisté, kteří prosazují evoluci člověka, nejsou objektivní ve své interpretaci fosilních nálezů?
Paleoantropolog Milford Wolpoff píše: „ Podle mého názoru ve vědě objektivita neexistuje. Dokonce i sbírání dat, která data zachovat a která ignorovat, zrcadlí vědcův světový názor“(2). Evolucionisté John Gribbin a Jeremy Cherfas přiznávají: „… musíme připustit, že historie paleontologie se nečte jako zářný příklad cesty za pravdou, zejména kdyby šlo o pravdu o původu člověka“(3). A dále připojují: „…víme dobře, že nic nemůže být vzdálenějšího realitě než populární obraz vědce jako věcného hledače pravdy“(4). Nakonec se zamysleme nad následujícím komentářem Rogera Lewina, autora knihy „Kosti sváru: Spory v hledání počátků člověka“:
„Jde o cosi jako obecný sebeklam (šířený většinou samotnými vědci), že při hledání objektivní pravdy jsou to právě poznatky, jež diktují závěry. Pokud by tomu tak bylo, pak by každý vědec mající stejné poznatky nutně dospěl k témuž závěru. Ale jak jsme již viděli a znovu a znovu uvidíme, k tomu často nedochází. Poznatky jsou velmi často překrucovány, aby se hodily k závěrům, které se nám líbí. A kardinální otázkou pak zůstává : „Co formuje priority vědce či skupiny vědců?“ , nikoli „Co je pravdivé?“ (5).
Vědci, jak evolucionisté tak kreacionisté, mají sklon interpretovat to, co ve světě vidí, svým vlastním zvláštním pohledem, který reprezentuje jejich pojetí věci, světový názor či ideologii. Pokud jejich „brýle“ mají evolucionistický ráz, pak budou obvykle zjištěná data překrucována tak, aby se hodila do tohoto preferovaného pojetí. Autor této studie si myslí, že je evolucionistické pojetí nesprávné, a že pouze biblický světový názor nám může pomoci správně pochopit náš původ. Podle Bible Bůh „stvořil z jednoho člověka všechno lidstvo“ (Skutky 17:26). Neexistuje tu prostor pro nějaké „opolidi“ jako předchůdce lidí, protože „od počátku stvoření je Bůh učinil jako muže a ženu“ (Marek 10:6). Proto všichni předpokládaní opolidé patří buď do rodu Homo (a jsou potomky Adama a Evy) či patří k vymřelým primátům. Ve svém článku užívá autor evoluční datování, aby umístil fosilie do evolučního kontextu: to však v žádném případě neznamená, že by snad s tímto datováním souhlasil.
Homo habilis (včetně Homo rudolfensis)
Lebeční kapacita se u Homo habilis pohybuje od těsně pod 500 kubických centimetrů (cm3) až ke zhruba 800 cm3 (6). Taxonomický zmatek je asi nejlepším výrazem pro vystižení současného statutu druhu habilis, jelikož se vášnivě debatuje o tom, zda fosilie mu připisované patří všechny jednomu druhu. Někteří experti rozštěpili tento druh do druhů dvou a vytvořili tak nový druh Homo rudolfensis, datovaný od zhruba 1,8 do 2,4 milionů let zpět (LZ). U druhu habilis zůstalo datování na zhruba 1,6 až 2,3 LZ – ovšem s méně exempláři zde zastoupenými (7). Aby se vnesl další zmatek do celého obrazu, Wood a Collard dokonce navrhli, aby druhy rudolfensis (reprezentovaný lebkou KNM-ER 1470) a habilis (lebka KNM-ER 1813) přešly z rodu Homo do Australopithecus (8). Tato myšlenka ovšem nenalezla širší podporu. To, že habilis je tvořen alespoň dvěma druhy, není také přijímáno jednomyslně – někteří evolucionisté tvrdí, že odchylky mezi exempláři habilis lze vysvětlit vnitrodruhovou variabilitou (9).

V diskusi kolem sporu o několika druzích v jednom poznamenal Wolpoff, že někteří vědci použili druh habilis „jako sáček na smetí“ (10). Tattersall a Schwartz popsali „status H. habilis jako všeobjímající druh – „odpadkový koš“, do něhož vědci pohodlně zametou celou heterogenní směsici fosilií“ (11). Homo habilis je často uváděn jako evoluční spojovací článek mezi australopiteky a Homo erectus, avšak dokonce i někteří evolucionisté připouštějí, že tato myšlenka je příliš jednoduchá. Podle Bernarda Wooda „Pokroky v technikách pro absolutní datování a nová měření fosilií samotných prokázaly, že prostý model jediné linie lidské evoluce, v němž Homo habilis následuje po australopitekovi a pak se vyvinul přes H. erectus v H. sapiens, je neudržitelný (12). Ze stanoviska kreacionisty je habilis neplatným taxonem – jde pouze o sbírku různých fosilií, jež můžeme zařadit buď mezi lidské (např. jako erectus) či mezi australopitekní primáty. Abych to ilustroval, uvádím níže několik příkladů.
Podle Wolpoffa (který označuje druh erectus za raného H. sapiens) je lebka KNM-ER 1813 z Koobi Fora, Keňa „spojována se spodinou lebeční a čelně-lícní oblastí, jež je tak podobná ranému H. sapiens, že autoři jako T.White zařazují ER 1813 do téhož taxonu (pro něho jde o H. erectus)“ (13). Wolpoff popisuje KNM – ER 1813 jako „velmi podobnou, vskutku nerozeznatelnou od raného H. sapiens svými zuby a čelně-lícní architekturou (kromě jejího úzkého středu tváří) – ale má mnohem menší objem mozku“ (14). Pokud je Whiteovo a Wolpoffovo hodnocení KNM-ER 1813 správné, pak tato lebka, s lebeční kapacitou pouhých zhruba 509 cm3 (15), patřila zřejmě velmi malému člověku.
Interpretace fosilní lebky KNM-ER 1470 z Koobi Fora, Keňa, jež má lebeční kapacitu asi 752 cm3, je problematická jak pro evolucionisty tak kreacionisty. Analýza kreacionisty Billa Mehlerta z roku 1999, jež se zaměřila na spornou rekonstrukci tváře lebky 1470, jej vedla k přesvědčení, že lebka „vypadá stále více jako štíhlý australopitek s poněkud větším mozkem (16). Avšak kreacionista Marvin Lubenow dlouho tvrdil, že jde o člověka, a ve své revidované a aktualizované knize „Kosti sváru“ nedávno prohlásil, že „srovnání naznačují, že lebka 1470 je modernější než jakákoli fosilie Homo erectus – dokonce i než materiál z Kow Swamp, jenž je starý pouze asi 10 000 let“ (17). Kreacionista Malcolm Bowden také tvrdil, že KNM-ER 1470 je „prostě lebka malého člověka“ (18). Ačkoli existují rozdíly mezi exempláři KNM-ER 1470 a KNM-ER 1813, mnoho z nich lze vysvětlit (podle evolucionisty Wolpoffa) takto: „Vezmeme-li v úvahu, že větší lebky a tváře s mohutným chrupem, vyvinutějším než je chrup psí (a jejich strukturální důsledky), exemplářů jako ER 1470, zračí změny v tělesné výšce“ (19). Ačkoli byla nalezena lebka gorily s touž lebeční kapacitou (752 cm3) (20) jako je kapacita KNM-ER 1470, posledně uvedená lebka je mnohem podobnější lebce lidské než lebce lidoopa.
Není cílem této práce podávat vyčerpávající přehled (případ po případu) fosilií, které evolucionisté zařazují do druhu habilis. Příkladem exempláře habilis, který velmi pravděpodobně patří k australopitekům (patrně druhu africanus) je však Stw 53 ze Sterkfontein, Jižní Afrika (21). Kuman a Clarke vypočítávají některé hlavní morfologické rysy Stw 53, jež podle jejich názoru opravňují k jeho zařazení do rodu Australopithecus, včetně zubů (jež jsou velmi dlouhé) a mozkovny, jež „je zpředu úzká a sevřená“ (22). Též obrázky počítačové tomografie ukazují, že v kostním labyrintu vnitřního ucha rozměry polokruhovitého kanálku lebky Stw 53 „svědčí ještě méně pro chůzi o dvou než je tomu u australopiteků“ (23).
Tohle by zřejmě vylučovalo lidský status tohoto exempláře (u něhož je obtížné odhadnout velikost mozku kvůli nekompletnosti nálezu) (24). Studie morfologie ušního labyrintu vedená Spoorem však shledala, že exemplář SK 847 ze Swartkrans, Jižní Afrika, spojovaný s habilis, má rozměry kanálku „podobné modernímu člověku“ (23). Jak zdůrazňují autoři a další vědci (25), SK 847 byla také spojována s erectus (např. Johanson ji srovnával s exemplářem erectus KNM-ER 3733) (26). Erectus je tedy nejpravděpodobnějším statusem pro SK 847, ale lebka je příliš fragmentární na to, aby bylo možno stanovit definitivní diagnózu. Ze spodiny lebeční odhadl Wolpoff lebeční kapacitu menší než 500 cm3 pro SK 847 (27), ale vzhledem k tomu, že chybí většina lebeční klenby, je tento odhad přinejlepším pouhé číslo, vzaté ze vzduchu.
Kromě lebek jsou často k habilis (či rudolfensis) řazeny moderní, lidsky vypadající stehenní kosti KNM-ER 1472 a KNM-ER 1481 – zejména proto, že byly nalezeny v téže lokalitě (Koobi Fora) jako lebka KNM-ER 1470. Jelikož však všechny pocházejí z různých sekcí vrstvy, nelze je navzájem spojovat (28). Analýza stehenní kosti KNM-ER 1481 ji identifikovala s erectus (29), což znamená, že se jedná o kost lidskou. Někteří evolucionisté však stále o ní raději mluví jako o habilis (30) – to však zřejmě proto, aby se neprokázala přítomnost erectus příliš brzy, jelikož kost datují asi 2 miliony let zpět. Je zřejmé, že v tomto fiktivním scénáři platí zákon, že čím dále v čase jde erectus, tím méně času zbývá habilis k tomu, aby se v něho vyvinul.

Nejdůležitějším z mimolebečních nálezů přičítaných habilis je částečná kostra OH 62 z olduvajské rokle, Tanzanie. Patřila jedinci, jenž byl v době nálezu „považován za tak malého či ještě menšího než jakýkoli známý fosilní hominid „(31). Analýzy proporcí končetin ukázaly pažně-stehenní index podobnější lidoopu než je tomu u kostry Lucy (afarensis). Tento výsledek je však podmíněn správností odhadů délky končetin (32). Většina z distální (od trupu vzdálenější) poloviny stehenní kosti OH 62 totiž chybí, takže její délku lze odhadovat pouze na základě srovnávání s jinými stehenními kostmi (33).
Jak se ukázalo nedávno, když Haeusler a McHenry použili jinou stehenní kost (OH 34) než je tradiční Lucy – AL 288-1 (často užívaná k odhadům), byl pažně-stehenní index pro exemplář OH 62 srovnatelný s indexem moderního člověka (34). Je tedy zřejmé, že jelikož proporce horních k dolním končetinám u OH 62 závisí na tom, která stehenní kost je použita pro srovnávání, přispívá tento údaj velmi málo k vyřešení taxonomického statutu OH 62. Avšak odhadovaná délka paže (vzhledem k relativně dlouhému předloktí) přesáhla délku paže moderních lidí a blížila se australopitekům a šimpanzům (35). Jak zdůrazňuje Lewin, byla to podobnost patra na lebce OH 62 s lebkou Stw 53, jež hrála rozhodující roli v připsání OH 62 druhu habilis (36). Jelikož je Stw 53 australopitekní lidoop (viz výše), bude jím OH 62 patrně také.
Nahoře je zobrazen výběr fosilních lebek. Exempláře ze Sangiranu a Ngandongu jsou řazeny jako Homo erectus. Hominid z jezer Willandra (WLH-50) a z Kow Swamp jako moderní lidé. Status lebky z Narmady je nejasný, jelikož není jisté, jak je „evolučně“ stará (věk obvykle udáván někde mezi 0,15 a 0,6 milionů let), je však řazena jako možný Homo heidelbergensis či archaický Homo sapiens. Studie prokázaly bližší příbuznost lebky moderního člověka WLH-50 s erectusem z Ngandongu, což je řadí mezi moderní svrchně pleistocenní lidi – Afričany či Levantince (79). Toto zjištění zpochybňuje myšlenku klasifikace Homo sapiens a Homo erectus jako samostatných druhů.
Homo erectus (včetně Homo ergaster)
Uvádíme zde, že Homo habilis tvoří fosilní exempláře, jež mohou být řazeny buď mezi vymřelé australopitekní lidoopy či mezi lidi. Některé z lidských exemplářů vykazují charakteristiky Homo erectus. Fosilie řazené mezi erectus považují evolucionisté za další vývojové stadium k moderním lidem, s habilis jako jejich údajným předkem. Použijeme-li prostý selský rozum, pak pokud by fosilie připisované erectusovi nebyly fosiliemi „opolidí“, ale plně lidské, pak se celý případ lidské evoluce v zásadě hroutí, jelikož existuje nepřeklenutelná morfologická propast mezi australopitekními lidoopy a lidským erectusem – mezi těmito druhy neexistují spojovací články.
Fosilní zbytky erectuse byly nalezeny v mnoha částech světa a evolucionisté je datují od asi 1,8 milionu let do cca méně než 100 000 let zpět (37). Fosilie H. erectus z Jávy, Indonésie, byly datovány možná až pouhých 27 000 let zpět, což je v evolučním schématu věcí skoro současnost (38). Multiregionální evolucionista Wolpoff hovoří o erectusovi jako o raném Homo sapiens, protože „nebyla nalezena jediná definice, jež odlišuje H. sapiens – definovaného tradičně jako následovníka H. erectus – od H. erectus ve všech regionech, kde se fosilie nalézají“ a „neexistuje zřetelný začátek H. sapiens po celou dobu, co uznáváme H. erectus“ (39).

Jiní přijímají pojetí se dvěma či více druhy, jež ve své nejjednodušší formě „odlišuje raně afrického H. erectus (tj. fosilie většinou z Koobi Fora a západní Turkany) jako H. ergaster, ale ponechává jiné africké H. erectus (např. OH 9) a asijské fosilie v rámci druhu H. erectus (sensu stricto)“ (40). V naší studii bude o erectusovi pojednáváno v širším smyslu, včetně fosilií z omezených definicí jak Homo erectus tak Homo ergaster. S průměrem asi 973 cm3 (41) je velikost mozku erectuse menší než je průměr asi 1 350 cm3 (42) pro žijící lidi. Rozsah lebeční kapacity pro erectuse (727 – 1 251 cm3) uváděný Rightmirem (43) je na spodní hranici nejširší definice rozsahu této kapacity pro moderního člověka (700 – 2 200 cm3) uváděného Molnarem (44). Molnar však neudal zdroj pro spodní limit (700 cm3)(45), a tak nejnižší ověřená velikost mozku pro normálního dospělého je asi hodnota zjištěná u Melanésana s lebeční kapacitou 790 cm3 (46).
Obrázek: Expozice v Muzeu člověka v San Diegu vystavuje repliku lebky KNM-ER 1470, v pozadí jsou dva modely „opolidí“.
Lebky připisované Rightmirem erectusovi zahrnovaly řadu Ngandong, řadu Zhoukoudian, OH 9, OH 12, Bouri, Trinil, řadu Sangiran, Dmanisi 2280, KNM-ER 3883, KNM-ER 3733, Buia, Gongwangling, Sale, Hexian, Ceprano a KNM-WT 15 000 (43). Rightmireova tabulka lebečních kapacit erectuse nezahrnovala lebku Dmanisi 2282 patřící „staršímu dítěti či mladému dospělému“ (cca 650 cm3) (47) a nejnovější lebku Dmanisi D 2700 (cca 600 cm3) (48), jejíž věk je stanoven jako ležící mezi juvenilním KNM-WT 15 000 a D 2282 (49). Ačkoli se jedná o exempláře dětské, nedalo by se očekávat, že by se lebeční kapacita těchto dvou exemplářů z Dmanisi (připisovaných erectusovi) nějak výrazně v dospělosti zvýšila. Další lebka z Dmanisi (D 2280) má odhadovanou lebeční kapacitu 775 cm3 (47). Je tu však také spodní čelist (D 2600) vykopaná roku 2000, jež byla popsána jako „enormní“ a „mnohem větší než jaká by se dobře hodila do kterékoli ze zatím objevených lebek“ (50).
Tyhle velké odchylky ve velikosti vedly k úvahám, že fosilie z Dmanisi (datované evolucionisty na 1,75 milionu let zpět) reprezentují více než jen jeden druh. Jelikož však byly nalezeny v téže stratigrafické vrstvě, je pravděpodobnější, že jsou všechny členy téže populace (51). Kamenné nástroje (52) nalezené v Dmanisi naznačují přítomnost lidí, a fosilní exempláře představují nejspíš zhotovitele těchto nástrojů. Že populace lidí z Dmanisi vykazuje velkou variabilitu ve velikostech lebek (a že velká mandibula D 2600 patřila patrně k mnohem větší lebce než jsou lebky ostatní), to jen potvrzuje známou pravdu, že také mezi žijícími lidmi existují enormní rozdíly ve velikosti lebek. Podobné či možná ještě větší rozdíly mezi velikostmi nálezů než u fosilií z Dmanisi jsou prý u lidských fosilií z ústí řeky Klasies (Jižní Afrika). Datovány od asi 120 000 do 90 000 let zpět, jsou považovány evolucionisty za „blízké modernímu člověku“. Variabilitu velikostí zde nejvíce reprezentují spodní čelisti, přičemž jednu z těchto mandibul (KRM 16424) popsal Klein jako „jednu z nejmenších čelistí dospělého člověka, jaké kdy byly zaznamenány“ (53).
Velikost mozku a inteligence
Podporuje rozdíl ve velikosti (sám o sobě) mezi mozkem běžného moderního člověka a průměrného erectuse evoluční myšlenku, že během údajného období evoluce hominidů se velikost mozku vyvíjela? Odpověď je záporná! Podle evolucionisty Hollowaye:
„Variační rozsah lebeční kapacity moderního Homo sapiens je kolem 1 000 cm3, a nemůžeme jasně doložit vztah mezi kapacitou a chováním. Uvedené číslo reprezentuje téměř celou šíři nárůstu kapacity od úrovně australopiteků po moderního člověka“ (54).
Lékař a antropolog John Relethford uznává, že „Ačkoli jejich mozek byl o něco menší než náš dnešní, Homo erectus měl v zásadě lidskou kostru od krku dolů, vyráběl složité kamenné nástroje, a možná užíval oheň“ (55). Schopnost vyrábět složité kamenné nástroje naznačuje, že menší mozek nebránil erectusovi v tom, aby měl lidskou inteligenci. Měli bychom si připomenout, že Anatole France, jehož mozek měl objem asi 1 000 cm3 (jen kousek nad průměrem erectuse), získal 1921 Nobelovu cenu za literaturu (56). Zamysleme se tedy nad tím, proč by se evoluce (kdyby to mohla činit) plahočila s vývojem většího mozku za velkou cenu, kdyby takový mozek neposkytoval žádnou hmatatelnou výhodu oproti mozku menšímu?
Evoluce je prý celá o přizpůsobovací hodnotě novinek v organizmu (z nichž může jen minimum znamenat přínos). Novinky v organizmu prý vznikly náhodně díky mimořádně nepravděpodobným genetickým mutacím. Z toho vyplývá, že nemá-li větší mozek žádnou zřejmou hodnotu pro adaptaci organizmu, pak by se jednoznačně nemohl vyvinout, i kdyby byla evoluce možná. Nikdo dosud neprokázal, jak by mohly samotné „přínosné“ genetické změny rozmnožit informační obsah genomu, jelikož tyto změny DNA v zásadě vyvolávají jen přeskupení a ztrátu informace (57). Mechanizmus pro vznik „přínosné“ evoluční změny zůstává tedy záhadnou „černou skříňkou“.
Mozek je téměř nekonečně složitý, a věřit, že jej nějaká neznámá přírodní síla hnala do stále se zvětšující velikosti (během údajné evoluce člověka) – aniž by se vlastně zvyšovala hodnota mozku pro adaptaci – znamená věřit v nulovou pravděpodobnost. Muselo se stát něco jiného, co by vysvětlovalo neuvěřitelnou variabilitu ve velikosti lidského mozku, a tím něčím byl inteligentní plán Stvořitele. Následující citát evolucionisty Hollowaye dokládá dilema takto zaměřených vědců:
„Jinak existuje určitá potíž s tvrzením, že přirozený výběr upřednostňoval větší mozky, jestliže nelze spojovat neurální struktury mozkové kůry s větší adaptabilitou v chování. Znamená to, že samotné jednotky, v nichž bychom vyjádřili pozvolný růst lebeční kapacity po celý pleistocén, centimetry krychlové, nemůžeme prokazatelně spojit s reálnými odlišnostmi v chování. Stav u moderního člověka, kde existuje variabilita téměř 1 000 cm3 beze změn v chování, jež lze analyzovat, nás varuje, že pokoušíme-li se vysvětlit nárůst lebeční kapacity během evoluce hominidů, musíme sáhnout k jinému parametru či parametrům“ (58).
Louis Leakey našel lebku Homo erectus OH 9 v roce 1960 v olduvajské rokli v Tanzanii. Je datována 1,2 milionu let zpět a má lebeční kapacitu 1067 cm3. Má mohutné nadočnicové oblouky. Snímky CT labyrintu kůstek vnitřního ucha tohoto exempláře naznačují morfologii moderního člověka, což svědčí pro chůzi o dvou. Snímek byl pořízen v Muzeu člověka v San Diegu.

To vše nezastavilo záplavu evolučních historek „tak nějak“ (o tom, jak se prý vyvinul náš velký lidský mozek), kulminujících v naprosté absurditě, že náš „enormní lidský mozek vytvořily paměťové stopy“ (59). Někdo se však může ptát, zda ty nejmenší exempláře erectuse měly mozky schopné lidské inteligence. Pokud bychom se řídili nedávnými fosilními nálezy klasifikovanými jako Homo floresiensis (více dále), pak musí být odpověď pozitivní, jelikož, slovy evolucionistky Kate Wongové, „Řekl by někdo, že stvořeníčko s lebkou velikosti grapefruitu mohlo mít poznávací schopnosti srovnatelné se schopnostmi anatomicky moderních lidí“ (60)?
Podle experta na erectuse Philipa Rightmirea z univerzity v Birminghamu: „Pokud byl Homo floresiensis schopen vyrábět složité nástroje, musíme říci, že pouhá velikost mozku mnoho neznamená“ (56). Měli bychom si pamatovat, jak prohlásil Holloway, že „Jeden kubický centimetr mozkové kůry šimpanze není totéž jako jeden cm3 kůry lidské. Ani není pravděpodobné, že může být nalezena jakákoli přiměřená poměřovací jednotka pro porovnávání mozků“ (61). Z toho plyne, že organizace neuronů je asi daleko důležitější než velikost mozku sama o sobě. Také platí (jak už jsme se zmínili výše), že neexistuje demonstrovatelná souvislost mezi lebeční kapacitou a chováním (včetně stupně inteligence), jak to napovídá následující výrok Clarka:
„Pokud bylo možné použít vhodné testy, nezjistil se v rámci jejich limitů žádný zřejmý vztah mezi velikostí mozku a inteligencí. Pro paleoantropologa je taková neexistence vztahu zvlášť znepokojivá, neboť to znamená, že nemá žádnou spolehlivou metodu ke zjištění mentální kapacity vymřelých hominidů pouhým stanovením kapacity lebeční“ (62).
Lokomoce a podlebeční kosti
CT záznamy labyrintu kůstek vnitřního ucha prokázaly u těch několika málo exemplářů erectuse, jež byly takto zkoumány (OH 9, Sangiran 2 a 4), morfologii moderního člověka, což svědčí pro chůzi o dvou (63). Jak jsme se již zmínili, uznávají evolucionisté, že celá kostra erectuse byla v podstatě lidská (55). První významná kompletní kostra erectuse (KNM-ER 1808) byla objevena roku 1973, byla datována 1,7 milionu let zpět, ale kvůli kostní chorobě, hypervitaminóze A, byla bezcenná s ohledem na poskytnutí jasného obrázku normální morfologie erectuse (64). Výška kostry ženy KNM-ER 1808 byla odhadnuta na 173 cm (65), a v době nálezu představovala zdaleka nejúplnější kostru erectuse – úplnější než všechny předchozí kostry erectuse dohromady(66).
Většina informací o anatomii kostry erectuse pochází z nálezu pozdějšího, na západě jezera Turkana v Keni roku 1984. Šlo o téměř kompletní kostru 1,68 m vysokého chlapce z Nariokotome (KNM-WT 15000) datovanou 1,6 milionu let zpět (67). Kostra je známa též jako chlapec z Turkany a zastánci teorie více druhů ji řadí pod Homo ergaster. Morfologie lebky chlapce z Nariokotome připomínala erectuse, ale Wolpoff popisuje kostru tohoto jedince jako „většinou moderní“ (68). Proporce končetin KNM-ER 15000, zvláště rameno-stehenní a pažní indexy, připomínaly moderní lidi (69). Podle Lewina naznačují informace získané z kostry chlapce z Nariokotome, že kostra erectuse „se podobá moderním lidem, ale je robustnější s mohutným svalstvem“, což „naznačuje rutinní těžkou fyzickou námahu“ (70). Lebeční kapacita chlapce, jemuž bylo asi 11 let, představovala v době jeho smrti asi 880 cm3, a odhad velikosti mozku v dospělosti je 909 cm3(71). Jak zdůraznil Mehlert, neexistuje způsob, jak stanovit s jistotou výšku tohoto jedince v dospělosti (72), ale jeden odhad pro KNM-ER 15000 hovoří o 185 cm (65).
Že se tak vysoký jedinec erectuse s moderní kostrou objevil v tak raném stadiu údajné evoluční historie erectuse, to je pro evolucionisty problémem. Kdyby byla evoluční teorie správná, pak bychom očekávali přechodnější postkraniální kostru, jež by svědčila více o přechodném stadiu mezi australopiteky a moderními lidmi, nikoli kostru již na úrovni moderního člověka. Existuje tedy velká morfologická mezera mezi erectusem a australopiteky; na taxon habilis (jak jsme probrali již dříve) pohlížíme jako na neplatný. V tom, co se jeví jako téměř zoufalá snaha dehumanizovat chlapce z Nariokotome, zdůrazňují někteří evolucionisté fakt, že páteřní kanál tohoto jedince byl užší než u moderních lidí (73).
Z toho vyvozují, že jeho nervová soustava nebyla dostatečně vyvinutá na to, aby zvládala celou škálu jemných dýchacích pohybů požadovaných pro plně lidskou řeč, a uzavírají, že v době, kdy chlapec žil, „se ještě plně nevyvinula řeč jak ji dnes chápeme“ (74). Avšak odhalení, že axiální kostra KNM-WT 15000 vykazovala značné abnormity (což napovídá, že tato kostra byla u hocha z Nariokotome abnormálně stavěna), činí neplatným jakoukoli takovouto námitku a může být objasněním pro úzkou stavbu páteřního kanálu (75). Tento závěr však někteří evolucionisté stále nepřijímají (76).
Charakteristické znaky lebek
O lebkách řazených pod erectuse se evolucionisté domnívají, že vykazují klíčové znaky, jež je odlišují od moderních lidí. Tyto znaky zahrnují: výrazné nadočnicové oblouky; nevýraznou bradu; velkou mandibulu; dopředu vystupující čelisti; ploché, ustupující čelo; dlouhou lebku s nízkou klenbou; vyvýšeninu na týlní kosti; relativně velké zuby; relativně velké obličejové kosti; a mozkovnu se silnými stěnami (77).
Velkým problémem pro evolucionisty je fakt, že mnoho (ne-li všechny) svrchu zmíněné znaky (jež prý odlišují erectuse od moderních lidí) se u moderních lidí vyskytují také. To dokládají nálezy u současných australských domorodců – výrazné nadočnicové oblouky lebky 3596 z Eustonu (78), a blízká příbuznost lebky moderního člověka z Austrálie, WLH – 50, s erectusem z Ngandongu, srovnávaným s moderními lidmi ze svrchního pleistocénu – Afričany a Levantinci (79). Podle Shreevea,
„Zatímco někteří z raných moderních lidí z Austrálie vypadají celkem jako lidé dnešní, jiní vykazují všechny známky robustnějšího druhu člověka, se silnými lebečními kostmi, mohutnými nadočnicovými oblouky a velkými zuby, dokonce i většími než jsou zuby druhu Homo erectus v některých jeho exemplářích“ (80).
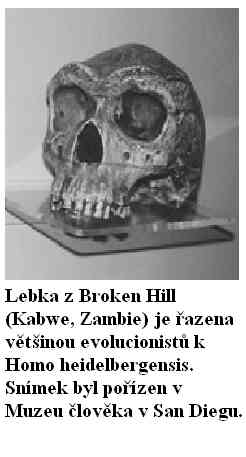
Obrázek: Lebka z Broken Hill (Kabwe, Zambie) je řazena většinou evolucionistů k Homo heidelbergensis. Snímek byl pořízen v Muzeu člověka v San Diegu.
Příklady dalších typických erectusovských znaků u moderních lidí, jako je plošší ustupující čelo a nevýrazná brada, můžeme vidět na fotografii žijícího australského domorodce publikované na sklonku viktoriánské éry, kdy v antropologii vládl úděsný rasismus (81). Domorodí Australané jsou lidští a „moderní“ jako kdokoli jiný, a tak nemohou být shora uvedené erectusovské rysy považovány za „primitivní“.
Stringer a Gamble, obhájci teorie, že moderní člověk má původ v Africe, napsali o přítomnosti erectusovských znaků u australských domorodců jako o zřejmě „jasných evolučních zvratech k dřívějšímu stadiu“ (82). Vzbudili tím rozzlobenou odpověď od skupiny konstatující, že „taková tvrzení a jejich důsledky jsou nešťastné“ (83).
I když ponecháme tyto spory stranou, je jisté, že uvedené tvrzení dobře ilustruje chameleonskou povahu evoluční teorie, jež se jeví dost tvárná na to, aby zastřešila téměř jakýkoli scénář. Je zřejmé, že neexistuje žádná přijatelná základna pro zavržení tvrzení, že fosilie erectuse jsou plně lidské – zavržení kvůli znakům lebky, jež někteří evolucionisté považují za „primitivní“. Kreacionisté přitom nejsou jediní, kdo protestuje proti úzké definici našeho lidského druhu. S ohledem na teorii původu člověka z Afriky vyjádřili evolucionisté z multiregionální názorové školy tuto obavu z příliš restriktivní definice druhu Homo sapiens:
„Myslíme si, že nešťastným aspektem debaty jsou (sic) definice Homo sapiens užívané některými zastánci teorie „Evy“. Jsou považovány za definice vylučující mnoho pleistocenních i mladších domorodých Australanů z našeho druhu (Wolpoff, 1986; P.Brown, 1990). Další zkoumání těchto jedinců i sbírek kosterních pozůstatků současných domorodců nás vede k odhadu, že tyhle definice moderního Homo sapiens vylučují zhruba 40 000 až 60 000 žijících domorodých Australanů. Cítíme, že se zde skrývá velké nebezpečí. Je povinností specialistů zajistit, aby jakákoli definice našeho druhu zahrnovala všechny žijící lidi. Budeme-li pak definovat člověka jako druh zahrnující minimálně všechny žijící lidi, pak mnohé z fosilií, o nichž zastánci „Evy“ prohlašují, že nezanechali potomky (včetně neandertálců) spadají pod Homo sapiens“ (84).

Pro účely analýzy byl erectus považován za zvláštní skupinu, ale je opravdu tak odlišný od jiných fosilních lidí? Například jsou fosilní exempláře připisovány erectusovi prostě kvůli morfologii svých lebek či existuje trend kategorizovat exempláře s menší lebeční kapacitou jako erectuse, a větší exempláře do jiných taxonů jako je Homo heidelbergensis či neandertálci? Než se naše diskuse přesune k další skupině fosilních lidí, ocitujme objevný komentář od evolucionisty Harryho Shapira:
„Zkoumáme-li však lebku klasického neandertálce (jichž máme nyní spoustu), nemůžeme se ubránit přesvědčení, že její základní anatomický tvar je zvětšenou a rozvinutou verzí lebky Homo erectus. Jako Homo erectus má hrbolkovitý výstupek v týlu, mohutné nadočnicové oblouky, relativně plochou hlavu, jež má směrem od týlu profil jako mansardová střecha. Ve své největší šířce je nízká, právě nad ušima, a typické je chybění vyčnívající brady“ (85).
Lebka neandertálce Gibraltar 1 byla nalezena ve Forbesově lomu, Gibraltar, před rokem 1848, a je datována někam mezi 45 000 a 70 000 let zpět. Říká se, že je to první nalezená lebka dospělého neandertálce, ale nebyla takto zařazována do té doby, než se našly originální fosilie neandertálce v jeskyni Feldhofer, Německo, v roce 1856.
Homo heidelbergensis (archaický Homo sapiens)
Homo heidelbergensis je kategorie vynalezená k vyplnění údajného taxonomického vakua mezi neandertálci a erectusem. Dříve byly tyto fosilie zařazovány pod archaického Homo sapiens (86). Evolucionista Shreeve popsal tento taxon jako „spíše cosi jako odpadkový koš, do něhož naházíte vše, co není ani jasný erectus, ani jasný moderní Homo sapiens“ (87). Mozkovny druhu heidelbergensis jsou popisovány jako „robustněji stavěné“ než mozkovny moderních lidí, a „mající některé, ne však všechny, znaky lebky erectuse, ale postrádající odvozené rysy lebky neandertálců“ (88).
Zda je účelné mít zvláštní druhy H. erectus a H. heidelbergensis je otázka, když například lebky erectuse série Ngandong (43) byly občas též řazeny pod archaický H. sapiens (tj. heidelbergensis) (89). Někteří evolucionisté dokonce obhajují začlenění fosilií Ngandong přímo k H. sapiens (90). Pakliže svrchu zmíněné lebky vykazují takovou zaměnitelnost v rámci rodu Homo, nemohou si evolucionisté stěžovat na to, že je kreacionisté házejí do jednoho pytle. Lebeční kapacita H. heidelbergensis sahá od 1 100 po 1 390 cm3 (průměr kolem 1 206 cm3), a jeho stáří se pohybuje mezi 200 000 a 700 000 lety zpět (91). Přehled exemplářů řazených pod H. heidelbergensis zahrnuje Dali, Broken Hill, Bodo, Arago, Jinniushan, Ndutu, Petralona, Steinheim, a Sima de los Huesos 4 a 5 (92). Shora zmíněné argumenty pro udělení lidského statutu erectusovi platí rovněž pro heidelbergensis, zvláště s ohledem na fakt, že na něj evolucionisté pohlížejí jako na „modernějšího“ než je erectus.
Homo antecessor
Tendence rozštěpovat rod Homo do více a více druhů je zřejmá i ze zařazení fragmentárních ostatků z Gran Dolina, Španělsko, do nového druhu, Homo antecessor. Ostatky zahrnují části obličejových kostí mladého jedince (ATD 6-69), jež mají „zcela moderní topografii tváře“ (93). Jistě by se byl našel prostor v „odpadkovém koši“ heidelbergensis i pro ostatky z Gran Dolina. Podezříváme zde vědce (jelikož evoluční stáří ostatků asi 780 000 let zpět je činí staršími než jsou jakékoli jiné fosilie druhu heidelbergensis v Evropě) ze snahy dát prostě jen těmto „nejstarším známým Evropanům“ nové jméno. To už je trochu moc, zejména poté, co vědci tvrdí, že antecessor „zaujímá klíčové postavení na rodokmenu člověka“ (94).
Homo neanderthalensis (neandertálec)
Evolucionisté věří, že neandertálci byli omezeni ve výskytu na Evropu, západní Asii a Střední východ mezi asi 30 000 a 150 000 lety zpět. Většina evolucionistů je považuje „za postranní větev lidského evolučního rodokmenu, jejímž osudem bylo vymřít“ (95). Relativně dlouhá historie objevů a přibývajících znalostí neandertálců byla vyprávěna již mnohokrát, i když většinou z hlediska evoluční teorie, a my se jí tu nebudeme znovu zabývat (96). Neandertálci měli relativně rozvinutou kulturu a pohřbívali své mrtvé (97). Každému, kdo není zaslepen evoluční předpojatostí, je již jen toto jasným důkazem toho, že neandertálci byli zcela lidští.
Exempláře klasických neandertálců zahrnují Neandertal, La Chapelle-aux-Saints, La Ferrassie I, Spy I, Le Moustier, Saccopastore II, Shanidar 1 a 5, Tabun a La Quina, zatímco „pokročilé“ exempláře zahrnují Spy II, Saccopastore I, Monte Circeo, ostatky Krapina, Shanidar 2 a některé z exemplářů Skuhl a Qafzeh (98). Neandertálci označení „klasický“ jsou považováni některými evolucionisty za „primitivnější“ (99).
Průměrná velikost mozku neandertálce zhruba 1 485 cm3 (rozsah: 1 245 – 1740 cm3) (100) je minimálně na stejné úrovni s moderními lidmi, ne-li o něco větší. Kromě velké lebeční kapacity uvádí Lubenow následující odlišné morfologické rysy neandertálce:
„(2) tvar lebky nízký, široký a protáhlý; (3) zátylek spíše špičatý, s hrbolem; (4) velké, mohutné nadočnicové oblouky; (5) nízké čelo; (6) velké, dlouhé obličejové kosti se středem tváře vyčnívajícím kupředu; (7) slabá, oblá brada; a postkraniální kostra robustní s velmi silnými kostmi“ (101).
Dalším charakteristickým znakem dospělých neandertálců je prostor za stoličkami, široké nozdry a velké zuby. Zatímco evolucionisté považují neandertálce za zvláštní druh, názorem kreacionisty je, že „erectus je pouhou zmenšenou kopií neandertálce a nejtypičtějším aspektem obou je tvar jejich lebky“ (103). Existují též neevoluční vysvětlení pro některé ze znaků neandertálců jako je tlak biomechanických sil, jenž ovlivnil morfologii jejich lebky (104).
Kromě toho demonstruje Jack Cuozzo ve své knize „Pohřbeni zaživa“ matoucí případy špatných rekonstrukcí některých exemplářů neandertálce. Na jednom příkladu ilustruje, jak byl poskládán exemplář Le Moustier tak, aby jeho čelist připomínala více opici než tomu bylo doopravdy (105), a na jiném příkladu předkládá Cuozzo důkazy o tom, jak vědci odřízli bradu exempláře La Quina 5, aby připomínal více opici (106).
Jak jsme již předeslali, mnohé z rysů, jež prý odlišují erectuse a neandertálce od moderních lidí, se u těchto lidí vyskytují také. Proto také věří zastánci multiregionálního pojetí lidské evoluce (na rozdíl od těch, kdo si myslí, že člověk pochází z Afriky), že Homo erectus, archaický Homo sapiens (heidelbergensis) a neandertálci „by měli být přeřazeni do jediného druhu, Homo sapiens, jenž je dále dělen pouze na rasy“, protože nejsou dostatečně odlišitelní od Homo sapiens (107). Zamysleme se nad následujícím výrokem zastánců multiregionální školy:
„Neandertálci mají mnohem větší nadočnicové oblouky než žijící Evropané, a ty se vždy táhnou přes celé čelo.Značné množství současných a žijících domorodých Australanů má velké nadočnicové oblouky přes celé čelo. Činí je to primitivnějšími než jsou Evropané? Činí to neandertálce moderními? „ (108).
Myslíte-li si, že jisté rysy lebky naznačují „primitivnější“ statut, pak svrchu zmíněné otázky znamenají skutečný problém.
Podle Stringera a Gamblea,
„Neandertálci nebyli opolidmi ani chybějícími spojovacími články – byli tak lidští jako my, ale představovali jiný druh lidí, druh s odlišnou směsicí primitivních a pokročilých znaků“ (109).
Tohle nám připadá jako přinejmenším matoucí prohlášení, protože jak může někdo říci, že neandertálci byli „tak lidští jako my“ a okamžitě nato připojit vymezení, že „představovali odlišný druh lidství“? Buďto tedy lidští byli nebo nebyli. Jak to trefně řekl Lubenow: „Problém neandertálců je v první řadě problémem evolucionistů. Stručně řečeno, evolucionisté nevědí, kde se tu neandertálci vzali či kam zmizeli“ (110). Pro kreacionisty byli neandertálci plně lidští. Neexistuje důvod, proč považovat některé znaky lebky za primitivnější než jiné, protože jsme všichni lidé, jeden jako druhý, bez ohledu na rozdíly v rysech, jež existují v rámci jediného druhu člověk, a protože opolidé nikdy neexistovali.
Homo floresiensis
Titulky médií koncem října 2004 jako „Zmizelá rasa lidských hobitů vykopána na indonéském ostrově“ (111) musely vyburcovat každého, kdo sleduje situaci s hominidy na této planetě. Protentokrát nebylo však neuvěřitelné pozdvižení v médiích (jež doprovázelo oznámení o dalším předpokládaném novém druhu hominidů, Homo floresiensis) přemrštěné, ačkoli udělení nového druhového jména těmto hobitům se zdá být trochu předčasné, jelikož (navzdory jejich malému vzrůstu) mohou pozůstatky docela dobře pocházet z potomků Adama (112). Tito hobité museli mít značné znalosti mořeplavby, když se dopravili na ostrov Flores, a vyspělé myšlení, „jak dosvědčuje způsob provedení kamenných artefaktů spojovaných s H. floresiensis v Liang Bua“ (113). Pokud nástroje opravdu patřily tomuto druhu (což se zdá být velmi pravděpodobné), pak tito lidé jistě měli lidskou inteligenci.
Nález fosilií z lidí velikosti hobita, již jak se zdá měli lidskou inteligenci, s jedním exemplářem (LB 1) datovaným evolucionisty asi 18 000 let zpět a vysokým asi jeden metr, s lebeční kapacitou zhruba 380 cm3 (114), zpochybňuje tvrzení o nejmenší možné velikosti mozku (alespoň 600 – 800 cm3) (115), který musí bytost mít, aby měla lidské mentální schopnosti. Lidé s mikrocefalními mozky (400 – 600 cm3) (116) stejně jako lidé malého vzrůstu (517 cm3) (117) mají též mozky pod touto dolní hranicí.
Průměrná velikost mozku šimpanzů je 383 cm3, orangutanů 404 cm3 a goril 504 cm3 (42). Mozek o kapacitě 380 cm3 u H. floresiensis z indonéského ostrova Flores je tedy velmi malý, je-li vůbec lidský. Avšak srovnáváme-li velikosti mozků, měli bychom též zohlednit výšku těla. To se děje, když vypočítáme hodnotu známou jako koeficient encefalizace (EQ) (118). Bylo-li tělo exempláře H. floresiensis LB 1 hubené a nízké, pak zjištěný EQ řadí tohoto jedince pohodlně do mantinelů třídy erectus (119). LB 1 je analogicky popisován takto: „Podle celkového tvaru lebky a zubů se bytost podobá nejspíše Homo erectusovi“ (120). Navzdory malé postavě a malé lebeční kapacitě měl LB 1 už jen málo společného s australopiteky. Podle autorů pojednání o H. floresiensis:
„…nemá velké postkaninní zuby, hlubokou tvář s vysunutou čelistí, a žvýkací schopnosti společné členům tohoto rodu. Naopak, obličejové a zubní proporce, postkraniální anatomie svědčící o chůzi o dvou běžné u lidí, a žvýkací aparát velmi podobný v relativní velikosti a funkci moderním lidem – to vše podporuje zařazení do rodu Homo. Pro toto zařazení mluví i údajná fylogeneze, jež nevylučuje tu a tam u H.erectus zakrňování“ (121).
„Rozdíly v anatomii kostry mohou prostě odrážet větší genetickou rozrůzněnost v rámci lidského druhu v minulosti.“
Peter Brown, paleoantropolog a první autor pojednání o floresiensis v časopise Nature, poznamenal v souvislosti s hobitovou malou lebkou, že „vnitřní struktura mozku – nervové dráhy – musely být spíše podobné lidským než opičím, aby mohl vyrábět takovéto nástroje“(122). Ještě pravděpodobnějším scénářem však je, že vnitřní uspořádání mozku floresiensis bylo prostě už lidské, jako tomu bylo u mozků mnoha jiných fosilních exemplářů s malými lebkami, zejména oněch připisovaných erectusovi.
Tým, který vykopal nález H. floresiensis, uvažuje o tom, že tito jedinci mohli být potomky erectuse z nedalekého ostrova Jávy (kde se prý vyskytoval před 1,6 milionem let) (123). Členové týmu soudí, že první hominidé připluvší na Flores „mohli být stejně velcí jako H. erectus a raný Homo a později zakrněli; nebo mohl přistát na Floresu ze sundského šelfu neznámý hominid s malým tělem i mozkem“ (119). Alternativním pohledem je názor, že floresiensis je „miniaturním človíčkem tvořícím část téže vrstvy post-bábelovské lidské rozmanité populace, jež zahrnuje i většího takzvaného Homo erectuse“ (124).
Patolog Maciej Henneberg z univerzity v Adelaide tvrdil, že jedinec LB 1 trpěl patologickou poruchou růstu zvanou sekundární mikrocefalie, a že „lebka hominida z Flores se velmi podobá 4 000 let staré mikrocefalní lebce moderního člověka nalezené na ostrově Kréta“ (125). A indonéský paleoantropolog Teuku Jacob prý řekl, že kosterní ostatky LB 1 pocházejí z „moderního člověka, Homo sapiens, jenž žil před asi 1 300 až 1 800 lety“ a byl „příslušníkem australomelanéské rasy, jež žila téměř na všech indonéských ostrovech“ (124). Týž paleoantropolog řekl, že lidé z Flores trpěli „mikrocefalií, jež smrštila jejich mozky na velikost mozků šimpanzích“ (126). Avšak čím více ostatků těchto drobných lidiček se nalézá, tím silněji se mluví proti verzi, že šlo o jedince nemocné. Máme zprávy o další spodní čelisti identické co do tvaru a velikosti s čelistí exempláře LB 1 (56).
Spory o hobita dost možná vyústí ve vytvoření dvou protichůdných táborů svářejících se nad problémem, trochu jako je tomu u trvající hádky mezi dvěma školami vyznávajícími učení o původu moderního člověka: školou „z Afriky“ a školou multiregionální. Pokud k tomu dojde, neočekávejte nějaké řešení brzy. Avšak i když se ukáže, že exempláře jsou patologické, naznačuje užívání složitých nástrojů (byť i „abnormálními“ lidmi), že byli docela inteligentní, a dokládá, že velikost mozku sama o sobě není tak důležitá jako neurální uspořádání.
Závěr
Homo habilis sestává z fosilních exemplářů, jež lze zařadit buď pod vymřelé australopitekní lidoopy či pod lidi (přičemž „lidští“ jedinci vykazují znaky Homo erectus). Bez klopotného úsilí zařazovat fosilie do evolučního schématu není důvod neuznat, že nálezy jako erectus a neandertálci patří do jediného rodu člověk. Rozdíly v anatomii koster mohou prostě jen odrážet větší genetickou diverzitu v rámci jediného rodu člověk v minulosti. V některých případech mohl život za zvlášť drsných podmínek či dokonce stravovací návyky ovlivnit vývoj kostry. Tlak zvláštních biomechanických sil stejně jako patologie (127) mohly též v některých případech ovlivňovat morfologii lebek a postkraniální kostry. V Bibli je též psáno, že lidé žili dříve déle, a že tato dlouhověkost po potopě pomalu ustupovala. Jelikož většina fosilií zřejmě patří raným popotopním lidem, je možné, že jejich přirozená doba dožití byla delší než je tomu u lidí dnešních. Je tedy nejasné, jaké důsledky by byla měla dlouhověkost, a možná i odlišná doba zrání kostry, na rysy fosilních lidí.
Odkazy na literaturu
1. Shermer, M., The shamans of scientism, Scientific American, 286(6):25, 2002.
2. Wolpoff, M.H., Paleoanthropology, Second Edition, McGraw-Hill, Boston, p.iv, 1999.
3. Gribbin, J., and Cherfas, J., The First Chimpanzee: In Search of Human Origins, Penguin Books, London, p.59, 2001.
4. Gribbin and Cherfas, ref.3, p.148.
5. Lewin, R., Bones of Contention: Controversies in the Search for Human Origins, Penguin Books, London, p.68, 1987.
6. Wood, B., and Richmond, B.G., Human evolution: taxonomy and paleobiology, J.Anatomy 196: 39, 2000.
7. Wood, B., and Collard, M., The human genus, Science 284: 66, 1999.
8. Wood, B., and Collard, M., The changing face of genus Homo, Evolutionary Anthropology, 8:204, 1999.
9. Miller, J.M.A., Craniofacial variation in Homo habilis: an analysis of the evidence for multiple species, American J. Physical Anthropology 112:103-128, 2000.
10. Wolpoff, ref.2, p.358.
11. Tattersall, I., and Schwartz, J.H., Extinct Humans, Westview Press, New York, p.111, 2001.
12. Wood, B., Origin and evolution of the genus Homo, Nature355:783, 1992.
13. Wolpoff, ref.2, pp. 369-370.
14. Wolpoff, ref.2, p.373.
15. Holloway, R.L., Problems of brain endocast interpretation and African hominid evolution; in: Jolly, C.J. (ed.), Early Hominids of Africa, Duckworth, London, p.387, 1978.
16. Mehlert, A.W., The rise and fall of skull KNM-ER 1470, Journal of Creation 13 (2):100, 1999.
17. Lubenow, M.L., Bones of Contention: A Creationist Assessment of Human Fossils, Revised and Updated, Baker Books, Grand Rapids, MI, p.328, 2004.
18. Bowden, M., Apemen: Fact or Fallacy? Second Enlarged Edition, Sovereign Publications, Bromley, Kent, p.200, 1981.
19. Wolpoff, ref.2, p.387.
20. Holloway, R.L., Cranial capacity, neural reorganization, and hominid evolution: a search for more suitable parameters, American Anthropologist, 68:113, 1966.
21. Berger, L.R., and Hilton-Barber, B., In the Footsteps of Eve: The Mystery of Human Origins, Adventure Press, National Geographic, Washington, D.C., pp.239-240, 2000.
22. Kuman, K., and Clarke, R.J., Stratigraphy, artefact industries and hominid associations for Strekfontein, Member 5, J.Human Evolution, 38:841, 2000.
23. Spoor, F., Wood, B., and Zonneveld, F., Implications of early homonid labyrinthine morphology for evolution of human bipedal locomotion, Nature 369: 648, 1994.
24. Tattersall and Schwartz, ref.11, p.113.
25. Tattersall and Schwartz, ref. 11, p.112; Wolpoff, ref.2, pp.383-384.
26. Johanson, D., and Edgar, B., From Lucy to Language, Simon and Schuster Editions, New York, p.184, 1996.
27. Wolpoff, ref.2, p.384.
28. Leakey, R.E.F., Evidence for an advanced Plio-Pleistocene hominid from East Rudolf, Kenya, Nature 242: 447-450, 1973.
29. Kennedy, G.E., A morphometric and taxonomic assessment of a hominine femur from the lower member, Koobi Fora, Lake Turkana, American J.Physical Anthropology 61: 433, 1983.
30. Trinkaus, E., Does KNM-ER 1481A establish Homo erectus at 2,0 myr BP? American J.Physical Anthropology 64: 137-139, 1984.
31. Johanson, D.C., et al., New partial skeleton of Homo habilis from Olduvai Gorge, Tanzania, Nature 327: 208, 1987.
32. Richmond, B.G., Aiello, L.C., and Wood, B.A., Early hominin limp proportions, J. Human Evolution, 43: 530-531, 2002.
33. Haeusler, M., and McHenry, H.M., Body proportions of Homo habilis reviewed, J. Human Evolution, 46: 433-437, 2004.
34. Haeusler and McHenry, ref.33, p.461.
35. Haeusler and McHenry, ref. 33, p.460.
36. Lewin, R., Human Evolution: An Illustrated Introduction, Fifth Edition , Blackwell Publishing, Malden, MA, p. 141, 2005.
37. Anton, S.C., Natural history of Homo erectus, Yearbook of Physical Anthropology 46: 128-132 (Supplement S 37 to the American Journal Physical Anthropology, Vol. 122), 2003.
38. Swisher, C.C. III, Rink, W.J., Anton, S.C., Schwarcz, H.P., Curtis, G.H., Suprijo, A., and Widiasmoro, et al., Latest Homo erectus of Java: potential contemporaneity with Homo sapiens in Southeast Asia , Science 274: 1870-1874, 1996.
39. Wolpoff, ref.2, pp. 396-397.
40. Anton, ref.37, p.154.
41. Rightmire, G.P., Brain size and encephalization in Early to Mid-Pleistocene Homo, American J.Physical Anthropology 124: 113, 2004.
42. Tobias, P.V., Evolution of the human brain; in: Gordon, E. (Ed.), Integrative Neuroscience, Harwood Academic Publishers, Amsterdam, p.39, 2000.
43. Rightmire, ref.41,p.110.
44. Molnar,S., Races, Types, and Ethnic Groups, Prentice-Hall Inc., NJ, p.57, 1975.
45. Foley, J., Creationist arguments: brain sizes, http://creationontheweb.com/redirect.php?http://www.talkorigins.org/faqs…, 17 November 2004.
46. Jue,D.S., Cranial capacity and endocranial casts, Journal of Creation4:56, 1990.
47. Gabunia, L., et al., Earliest Pleistocene hominid cranial remains from Dmanisi, Republic of Georgia: taxonomy, geological setting and age, Science 288: 1019, 2000. .
48. Vekua, A., et al., A new skull of early Homo from Dmanisi, Georgia, Science 297: 88, 2002.
49. Vekua, ref.48, p.86.
50. Wong, K., Stranger in a New Land, Scientific American 289 (5): 56, 2003.
51. Wong, ref.50, p.59.
52. Gabunia, L., Anton, S.C., Lordkipanidze, D., Vekua, A., Justus, A., and Swisher III, C.C., Dmanisi and dispersal, Evolutionary Anthropology 10:164, 2001.
53. Klein, R.K., and Edgar,B., The Dawn of Human Culture, John Wiley and Sons Inc., New York, pp. 227-229, 2002.
54. Holloway, ref. 20, p.103.
55. Relethford, J.H., Reflections of Our Past: How Human History is Revealed in our Genes, Westview Press, Boulder, CO, p.46, 2003.
56. Wong, K., The littlest human, Scientific American, 292(2):48, 2005.
57. Sarfati, J., Refuting Evolution 2, Master Books, Green Forest, AR, pp. 56, 101-108, 2002.
58. Holloway, ref.20, p.115.
59. Blackmore, S., The Meme Machine, Oxford University Press, Oxford, p.81, 1999; Pro kritické zhodnocení této knihy, viz: Line, P., Unleashing the meme: is this the end of our existence? Creation Matters, 7(2), 2002.
60. Wong, ref.56, p.42.
61. Holloway, ref.20, p.108.
62. Clark, W.E. Le Gros, Bone of contention; in: Korn, N., and Thompson, F.W. (Eds.), Readings in Physical Anthropology, Holt, Reinehart and Winston Inc., New York; citováno v: Jue, ref.46, p.57.
63. Spoor et al.,ref.23, pp. 645-648.
64. Walker, A., and Shipman, P., The Wisdom of Bones: In Search of Human Origins, Phoenix, London, pp.128-134, 1996.
65. Wolpoff, ref.2, p.408.
66. Walker and Shipman, ref.64, p.157.
67. Brown, F., Harris, J., Leakey, R., and Walker, A., Early Homo erectus skeleton from west Lake Turkana, Kenya, Nature 316: 788, 1985.
68. Wolpoff, ref.2, p.402-403.
69. Richmond et al., ref.32, pp. 534-542.
70. Lewin, ref.36, p.163.
71. Walker and Shipman, ref.64, p.184.
72. Mehlert, A.W., Homo erectus „to“ modern man: evolution or human variability? Journal of Creation 8 (1): 107, 1994.
73. MacLarnon, A., and Hewitt, G., Increased breathing control: another factor in the evolution of human language, Evolutionary Anthropology, 13: 186-187, 2004.
74. Stringer, C., and McKie, R., African Exodus: The Origins of Modern Humanity, Jonathan Cape, London, p.30, 1996.
75. Latimer, B., and Ohman, J.C., Axial dysplasia in Homo erectus, J.Human Evolution 40: A12, 2001.
76. MacLarnon and Hewitt, ref.73, pp. 194-195.
77. Lubenow, ref. 17, p.122; Lewin, ref. 36, p.163; Klein, R.G., The Human Career, Second Edition, The University of Chicago Press, Chicago, pp. 283-289, 1999.
78. Hawks, J., Oh, S., Hunley, K., Dobson, S., Cabana, G., Dayalu, P., and Wolpoff, M.H., An Australian test of the recent African origin theory using the WLH-50 calvarium, J.Human Evolution, 39: 17, 2000.
79. Hawks, et al., ref.78, pp. 1-22; Wolpoff, M.H., Hawks,J., Frayer, D.W., and Hunley,K., Modern human ancestry at the peripheries: a test of the replacement theory, Science 291: 293-297, 2001.
80. Shreeve, J., The Neandertal Enigma, William Morrow and Company Inc., New York, p.100, 1995.
81. Hutchinson, H.N., Gregory, J.W., and Lydekker, R., The Living Races of Mankind, Hazell, Watson and Viney Ltd., London, pp. 60, nedatováno, ale pozdně viktoriánské.
82. Stringer, C.B., and Andrews, P., Genetic and fossil evidence for the origin of modern humans, Science 239: 1267, 1988.
83. Wolpoff, M.H., Spuhler, J.N., Smith, F.H., Radowcic, J., Pope, G., Frayer, D.W., Eckhardt, R., and Clarke, G., Modern human origins, Science 241: 772, 1988; viz též : Shreeve, ref. 80, p.109.
84. Thorne, A., and Wolpoff, M.H., Conflict over modern human origins, Search 22(5): 177, 1991.
85. Shapiro, H.L., Peking Man, George Allen et Unwin Ltd., London, p.125, 1974.
86. Wood and Collard, ref.8, p.197; Lubenow, ref.17, p.199.
87. Shreeve, ref.80, p.82.
88. Wood and Collard, ref. 8, p.197.
89. Brauer, G., and Mbua, E., Homo erectus features used in cladistics and their variability in Asian and African hominids, J.Human Evolution, 22:82, 1992.
90. Hawks et al., ref. 78, p.21.
91. Rightmire, ref. 41, pp.112-115.
92. Rightmire, ref. 41, p.112.
93. Bermudez de Castro, J.M., Arsuaga, J.L., Carbonell, E., Rosas, A., Martinez, I., and Mosquera, M., A hominid from the lower Pleistocene of Atapuerca, Spain: possible ancestor to Neandertals and modern humans, Science 276: 1392, 1997.
94. Gibbons, A., A new face for human ancestors, Science 276: 1331, 1997.
95. Lewin, ref. 36, pp. 179, 185.
96. Viz například: Tattersall, I., The Last Neanderthal: The Rise, Success, and Mysterious Extinction of Our Closest Human Relatives, Macmillan, New York, pp. 74- 119, 1995.
97. Lubenow, ref. 17, pp. 236-257.
98. Beasley, G.J., A possible creationist interpretation of archaic fossil human remains, Journal of Creation 6(2): 144, 1992.
99. Beasley, ref. 98, p.147.
100. Holloway, R.L., The poor brain of Homo sapiens neanderthalensis: see what you please… in: Delson, E. (Ed.), Ancestors: The Hard Evidence, Alan R.Liss, New York, pp. 320-321, 1985.
101. Lubenow, ref. 17, p.77.
102. Lewin, ref.36, p.179.
103. Lubenow, ref. 17, p. 258.
104. Lubenow, ref. 17, pp. 82-85.
105. Cuozzo, J., Buried Alive: The Startling Truth about Neanderthal Man, Master Books, Green Forest, AR, pp. 166-167, 1998.
106. Cuozzo, ref.105, pp. 41-42.
107. Shipman, P., On the origin of races, New Scientist 137: 34, 1993.
108. Wolpoff, M., and Caspari, R., Race and Human Evolution, Simon et Schuster, New York, pp. 331-332, 1997.
109. Stringer, C., and Gamble, C., In Search of the Neanderthals: Solving the Puzzle of Human Origins, Thames and Hudson, London, p.219, 1993.
110. Lubenow, ref. 17, p. 80.
111. Cauchi, S., Lost race of human „hobbits“ unearthed on Indonesian island, The Age, p.1, 28 October, 2004.
112. Wieland, C., Soggy dwarf bones, www.creationontheweb.com/content/view/3211, 28 October 2004.
113. Morwood, M.J., et al., Archaeology and age of a new hominin from Flores in eastern Indonesia, Nature 431: 1091, 2004.
114. Brown, P., Sutikna, T., Morwood, M.J., Soejono, R.P., Jatmiko, E., Saptomo,W., and Due, R.A., A new small-bodied hominin from the late Pleistocene of Flores, Indonesia, Nature 431: 1055-1061, 2004.
115. Leakey, L.S.B., Tobias, P.V., and Napier, J.R., A new species of the genus Homo from Olduvai Gorge, Nature 202: 7, 1964.
116. Holloway, ref.20, p.107.
117. Passingham, R.E., and Ettlinger, G., A comparison of cortical functions in man and the other primates, International Review of Neurobiology, 16: 257, 1974.
118. Aiello, L. and Dean, C., An Introduction to Human Evolutionary Anatomy, Academic Press, London, pp.189-191, 1990.
119. Brown et al., ref.114, p.1060.
120. Knight, W., and Nowak, R., Meet our new human relatives, New Scientist, 184 (2471): 8, 2004.
121. Brown et al., ref.114, pp.1060-1061.
122. Knight and Nowak, ref.120, p.9.
123. Dalton, R., Little lady of Flores forces rethink of human evolution, Nature 431: 1029, 2004.
124. Wieland, C., Hobbling the Hobbit, www.creationontheweb.com/content/view/3219, 8 November 2004.
125. Balter, M., Skeptics question whether Flores hominid is a new species, Science, 306: 1116, 2004.
126. Aglionby, J., and McKie, R., Hobbit folk „were just sick humans“, observer.guardian.co.uk/international/story/0,6903,1345519,00.html, 9 November 2004
127. Beasley, ref.98, pp.142-143.