
Část II.: Hominidi nepatřící do rodu Homo
(Z creation.com/ přeložil M. T. – 02/2007. První část naleznete zde – Část I. Rod Homo.
Cílem tohoto přehledu je podat kritickou analýzu zkamenělin údajných opolidí, které neřadíme do rodu Homo. Podrobnosti o všech těchto živočiších aspirujících na status hominidů probereme zhruba v tom pořadí, jak je stanovili evolucionisté, přičemž začneme tím nejstarším. Morfologie koster těchto údajných opolidí svědčí o tom, že se jedná o vymřelé lidoopy nemající nic společného s evolucí člověka.
Údajní opolidé patří do kategorie zkamenělin známé jako hominidi, což je skupina zahrnující žijící lidi, jejich předky i veškeré pitekoidní (opicím podobné) tvory považované za bližší lidem než šimpanzům. Někteří evolucionisté označují nyní tuhle skupinu jako homininy (1). Termín hominid považují kreacionisté za absurdní, protože podle nich byli hominidé buď lidmi nebo lidoopy, nikoli něčím přechodným. Zmíněný termín zde tedy užíváme jen pro účely kategorizace zkamenělin a v žádném případě netvrdíme, že takoví tvorové skutečně existovali. Podle evolucionistů je rozhodujícím okamžikem pro zařazení živočicha mezi hominidy údajné rozdělení vývojových větví od společného hmyzožravého prapředka na linii směřující k lidem a linii směřující k lidoopům před zhruba šesti miliony let.
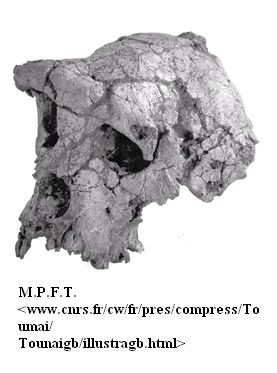
Obrázek: Údaje o Sahelanthropus tchadensis, známém nejvíce svou lebkou (TM 266-01-060-1), přezdívaném Toumai, byly zveřejněny v červenci 2002 a označeny jako nález nejstaršího známého hominida; přitáhlo to mimořádnou pozornost vědců. Přeřazení tohoto tvora některými experty mezi lidoopy však už takovou pozornost médií nevyvolalo.
V minulosti byl stanoven kalibrační bod rozštěpu lidských a šimpanzích linií pomocí molekulárních hodin na dobu před 5 miliony let (Ma) – jako pomocný kalibrační bod přitom posloužil údajný rozštěp mezi opicemi a lidoopy Starého světa před 30 Ma stanovený konvenčním radiometrickým datováním fosilií (2). Ke zmíněnému rozštěpu teď podle evolucionistů došlo před 5-7 Ma. Radiometrická datování (3) i molekulární hodiny (4) jsou založeny na neověřených a problematických předpokladech. Udáváme-li v tomto článku takováto evoluční stáří zkamenělin, činíme tak proto, abychom je zařadili do evolucionistického kontextu a v žádném případě to neznamená, že takto určená stáří považujeme za odůvodněná.
Za australopiteky jsou považováni členové rodu Australopithecus a dříve se jednalo o jediný rod hominidů kromě rodu Homo. V poslední době bylo však mnoho hominidů přeřazeno z rodu Australopithecus do nových taxonů jako třeba rodů Ardipithecus, Sahelanthropus, Orrorin a Kenyanthropus. Z kreacionistického hlediska bychom si měli uvědomit, že evolucionistický druh /species/ není totožný s druhem biblickým (5).
Sahelanthropus tchadensis
Je-li publicita obklopující oznámení nálezu fosilie něčím, co by nás mělo zajímat, pak je Sahelanthropus tchadensis nejlepším adeptem na titul nejstaršího hominida, jelikož ho provázel rozruch větší než jiné fosilie opolidí proslulé tím, že při svém objevení nadělaly rovněž spoustu „pozdvižení“. Lebka Sahelanthropa (TM 266-01-060-1, přezdívaná Toumai) nalezená v Čadu ve střední Africe figurovala na obálce časopisu Nature z 11. července 2002 s nápisem „Nejstarší známý hominid“. Tuhle jedinou lebku i další zkameněliny zmíněného tvora (úlomky spodních čelistí) datovali badatelé s využitím doprovodné fauny do doby mezi 6 a 7 Ma, přičemž lebka měla podle nich objem mozkové dutiny zhruba mezi 320 a 380 cm3 (6). Podivné na lebce Toumai je to, že zezadu vypadá jako lebka šimpanze, zepředu však podle jednoho experta připomíná vyspělého australopiteka (7).
Podle Bernarda Wooda „by neměl mít hominid Toumaiova předpokládaného stáří tvář menší jak do třetiny svého geologického věku“ (7). Pokládají-li tedy evolucionisté Sahelanthropa za nejstaršího vůdčího hominida, pak to vede k oprávněným pochybám i o statutu všech dalších hominidů s „primitivnějším“ vzhledem (7) jako předků člověka a eo ipso by to rovněž zpochybňovalo status většiny geologicky mladších australopiteků.
Přitom skoro ještě ani neodezněl mediální povyk kolem nálezu Toumaie a další evolucionisté včetně Brigitte Senutové z Přírodopisného muzea v Paříži vyjádřili pochybnosti o statutu Toumaie coby hominida. V jednom interview zmíněná dáma prohlásila, že se kloní „k názoru, že se v tomto případě jedná o lebku gorilí samice“. Takové úvahy jsou v příkrém rozporu s dřívějším nadšeným tvrzením harvardského paleontologa Daniela Liebermana o tom, že Toumai je „nejdůležitější zkamenělinou objevenou za dobu existence lidstva“ (9).
Tým vedený Wolpoffem zase namítl, že Toumaiovy mohutné nadočnicové oblouky větší než u kteréhokoli australopiteka či žijícího lidoopa jsou spíše mechanickou odpovědí na zátěž (10) a nikoli známkou morfologické podobnosti s rodem Homo. Zkoumání jizev po úponech šíjových svalů vzadu na lebce pak dovedlo zmíněný tým k závěru, že Toumai byl kvadrupedem (tj. chodil po čtyřech, nikoli po dvou) (11). Poté, co takto Toumaie prozkoumali, vyjádřili členové týmu přesvědčení, „že Sahelanthropus byl lidoopem“ (12). Dalším údajným hominidním znakem Toumaie jsou ve srovnání se žijícími lidoopy menší špičáky (13). Ačkoli se Sahelanthropus v některých znacích od žijících goril liší, je zde i mnoho podobností, a podle Matthewa Murdocka je docela dobře možné, že je Sahelanthropus příbuzný žijícím gorilám v tom smyslu, že jsou oba zástupci téhož biblického druhu (14).
Platí-li tahle premisa, pak Toumaiovy menší špičáky svědčí prostě o větší proměnlivosti těchto zubů u tohoto druhu lidoopů v minulosti; genetická informace kódující malé špičáky se pak vytratila. Nedávno se objevily i námitky toho druhu, že jedna ze stoliček v mandibule byla při rekonstrukci lebky vlepena na nesprávné místo a že o jednom řezáku nalezeném při vykopávkách se v původním článku časopisu Nature nepsalo (15). Autoři této kritiky argumentovali, že zkameněliny připisované Sahelanthropovi „jsou přece stále ještě předmětem diskuze o tom, zda představují hominida či lidoopa. Z tohoto důvodu je třeba poskytnout vědecké obci úplné a přesné údaje o nalezených fosiliích“ (16).
Ovšem jak tomu často bývá v paleoantropologii v situaci, kdy někdo zpochybní práci vědeckých kapacit, setkali se i zde jejich kritičtí kolegové s prudkou nesnášenlivou reakcí (17). Rovněž bylo nasnadě, že taková „degradace“ Sahelanthropa na lidoopa se jako obvykle v podobných sporech nesetká zdaleka s takovou pozorností médií, s jakou uvítala právě média původně povýšení zmíněného tvora mezi opolidi.
Orrorin tugenensis
Počátkem roku 2001 oznámil tým badatelů vedený Brigitte Senutovou a Martinem Pickfordem nález zkamenělin tvora Orrorin tugenensis označeného za možného nejstaršího hominida žijícího prý před zhruba 6 miliony let (jeho fosilizované kosti se našly v keňské Tugenské vysočině). Nález tvořilo 13 úlomků včetně částí stehenních kostí, několika zubů a kousků dolní čelisti (19).
Ze zmíněných ostatků není tedy možné zjistit, jak vlastně hlava tohoto tvora vypadala. Jediným důvodem pro zařazení Orrorina mezi rané hominidy i pro postulování jeho bipedie je proto fakt, že jeho fosilie pocházejí z toho správného evolučního období. Bipedie je evolucionisty považována za důležitý důkaz opravňující k udělení statutu hominida. Orrorin prý kráčel po dvou nohou vzhledem ke svému údajnému hominoidnímu femuru (stehenní kosti) s dlouhým stehenním krčkem a jamkou na jeho hřbetu vyskytující se u lidí, nikoli však u šimpanzů (20).
Tuto jamku v místě, kde se na kost upíná sval musculus obturatorius externus, nalézáme však i u nebipedů (21), a podle jednoho experta skeny napříč Orrorinova femuru ukazují na šimpanzoidní rozložení kortikální kosti (20).
(musculus obturatorius externus = sval v oblasti stehna. Začíná z vnější plochy membrana obturatoria a okolí, upíná se na femur do fossa trochanterica. Funkcí je zevní rotace stehna, dále addukce a flexe. Pozn. překl.)
(kortikální kost = kompaktní kost, tvořena převážně kompaktou, tj. druhem kostní tkáně vytvořené z haverských systémů. Haverský systém – osteon = struktura v kostní tkáni. Kostní hmota je zde uspořádána do longitudinálních lamel koncentricky utvářených okolo Haversových kanálů, jimiž procházejí cévy a nervy. Clopton Havers, 1650-1702, anglický anatom a lékař. Pozn. překl.)
Dalším argumentem zastánců jeho hominidního statutu je fakt, že hlavice stehenní kosti Orrorina je relativně mohutnější než u Lucy a že takový femur prý pomáhá rozložit tlaky vyvolávané bipedií (22). Zmíněný tým přišel také se spornou teorií o tom, že u Orrorina se bipedie vyvinula nezávisle na Lucy i jiných druzích australopiteků, které zařadil do vymřelé vedlejší větve čeledi Hominidae (22).
Spory kolem Orrorina se rozhořely opět v září 2004 po zveřejnění výsledků naskenování jeho femuru pomocí počítačové tomografie (CT), které vedly vědce k závěrům o nelidoopí, pozdně hominidní distribuci kortikální kosti na spoji střední části femuru a jeho krčku svědčícím o bipedální lokomoci (23). Ann Gibbonsová se ve svém referátu o těchto nejnovějších objevech vyjádřila takto:
„Senutová přišla s názorem, že Orrorinovo držení těla při chůzi bylo antropoidnější než u 2 až 4 miliony let starých australopiteků. Bylo-li tomu opravdu tak, pak by to vyřazovalo australopiteky z přímé linie předků člověka – dramatická revize naší prehistorie“ (24).
Paleoantropolog Tim White vyjádřil nesouhlas s tímto hodnocením nálezů podle „způsobu chůze“ a napadl nejnověji publikované závěry vědců s tím, že „rozlišovací schopnost CT skenů je tak mizerná, že na nich není jasně patrná struktura kostní hmoty“ (24). White rovněž „nazval degradaci australopiteků provedenou Senutovou ‚kreacionistickým názorem‘, protože naznačuje, že Orrorinův femur byl před 6 miliony let docela moderní a nevyvíjel se tedy po krůčcích“ (24). Zdá se, že i evolucionisté odchylující se ve svém výkladu dějin člověka od pravověrné evoluce riskují odmítavé reakce.
Podle Davida Beguna z torontské univerzity jsou důkazy o bipedii u Orrorina sporné (21). Zmíněný vědec napsal, že z nalezených zkamenělých úlomků kostí Orrorina nelze zjistit, zda šlo „o předka lidí, šimpanzů, o společného předka obou linií či o pouhého příslušníka vymřelé vedlejší vývojové linie“ (19).
Ardipithecus kadabba
Pár měsíců po oznámení nálezu druhu Orrorin tugenensis byl v časopisu Nature popsán další kandidát na raného hominida ze středního toku Awaše v Etiopii zvaný Ardipithecus ramidus kadabba, starý asi 5.2 až 5.8 milionů let (25).
(Awaš = řeka v Etiopii; délka cca 900 km. Archeologické lokality na jejím toku, kde byly objeveny kosterní pozůstatky předchůdců člověka, jsou součástí světového kulturního dědictví UNESCO. Pozn. překl.)
Zmíněný nový taxon byl roku 2004 po nálezu dalších fosilních zubů povýšen z poddruhu na samostatný druh, Ardipithecus kadabba (26). Stejně jako v případě Orrorina tvořilo i u tohoto druhu nalezené zbytky několik úlomků lebečních kostí, pár zubů a úlomky čelisti, takže není ani známo, jak vlastně hlava těchto tvorů vypadala.
Předpokládá se, že nalezený kadabba byl bipední; vědci tak usuzují ze znaků jediné kosti prstu u nohy (proximálního článku prstu = bližšího trupu, např. palce či ukazováčku), zejména z nasměrování proximální plochy tohoto kloubu k zádům (27). Vědci tvrdí, že plocha kloubu zmíněného prstu je humanoidně zvednuta vzhůru, zatímco u šimpanzů se ohýbá dolů; to je podle nich důkazem toho, že kadabba „našlapoval při chůzi na špičky“ tak jako lidé (28). Jak však zdůraznil Begun,
„se stejným postavením kloubu se setkáváme u hominida z mladšího miocénu Sivapitheka, který rozhodně po dvou nechodil; délka i zakřivení nalezené kosti pak nápadně připomínají znaky kostí šimpanzích či bonobích“ (29).
Zarážející je také ta skutečnost, že zmíněný článek prstu nohy je prý o několik set tisíc let mladší než ostatní nalezené fosilie a že se našel v lokalitě vzdálené od nich 16 km; to způsobilo, že ani slavný lovec hominidů Donald Johanson si není jist, jaký vztah má uvedená osamělá kost k ostatním fosiliím (30).
Fakt, že máme k dispozici pouze několik málo (jedenáct) fosilních zbytečků patřících nejméně pěti různým jedincům a nalezených na pěti různých místech (25), zpochybňuje jejich zařazení k témuž poddruhu a později, po nalezení několika dalších zubů (26), vytvoření nového druhu. Podle Baltera a Gibbonse „Tvrdí týmy, které našly Orrorina a Ardipitheka, že zkameněliny nalezené kolegy by mohly představovat předky šimpanzů či jiných lidoopů spíše než předky raných lidí či jejich blízké příbuzné“ (31). Snad mají oba týmy trochu pravdu, jelikož všechno nasvědčuje tomu, že oba hominidi byli pouhými lidoopy.
Ardipithecus ramidus
Roku 1994 ohlásil tým vedený Timem Whitem nález tvora Australopithecus ramidus na lokalitě Aramis v Etiopii, který v oné době představoval se svým stářím 4,4 milionu let nejstarší relevantní nález „hominida“ (32). Po zhruba osmi měsících překlasifikovali autoři nálezu v corrigendu ke své původní zprávě fosilie do zcela nového rodu, Ardipithecus, a pojmenovali je novým druhovým názvem Ardipithecus ramidus (33). K původnímu článku o zmíněném nálezu připojil časopis Nature z 22. září 1994 na své obálce fotografii představující úlomek čelisti Ardipitheka ramida, do níž byl zasazen jeden zub, to vše s nadpisem „Nejstarší hominidi“. Byli bychom tedy očekávali nález značné důležitosti, zatím však ony zkameněliny sestávající ze zbytečků kosti týlní, zubů a lebky vykazují, řečeno s autory nálezu, „spoustu znaků obvykle patrných na moderních lidoopech“ (34). Přesto zmínění paleoantropologové tvrdí, že jde o hominida, a opírají tento svůj závěr o to, že nalezený špičák vypadá na rozdíl od špičáků lidoopů spíše jako řezák, a o fakt, že foramen magnum (otvor v kosti týlní, kterým prochází prodloužená mícha) je posunut více dopředu; tito vědci mají také za to, že „Přítomnost uvedených znaků u nálezu z Aramis by mohla svědčit o bipedii, ačkoli se taková hypotéza samozřejmě musí ještě prověřit“ (35).
Ani v samotné evolucionistické komunitě nejsou pak všichni přesvědčeni o tom, že ramidus je hominidem. Podle Petera Andrewse z Přírodopisného muzea v Londýně je tenká vrstva skloviny na ramidových zubech „přece jen silnější než sklovina, kterou bychom očekávali u fosilního šimpanze“ ale znaky kosti pažní „svědčí o chůzi po kloubcích prstů jako u šimpanzů“ (36). Nedávno se však v Afaru v Etiopii našly další zkamenělé zbytečky (kousky čelistí, zubů, kosti prstů nohou i rukou) připisované nejméně devíti jedincům ramida a zúčastnění badatelé se vrátili znovu k úvahám o bipedii opírajícím se o dorzální orientaci proximální plochy kloubů proximálních článků prstů nohou (tedy o stočení palce či ukazováčku na noze směrem dozadu), znak pozorovaný i u kadabby (37).
(Afar = lokalita ve východní Etiopii; jedno z nalezišť kosterních pozůstatků hominidů náležejících do rodu Australopithecus (např. tzv. Lucy) a mnoha kosterních zbytků fosilní fauny. Datování asi 2-6 milionů let. Na základě zdejších nálezů byl popsán gracilní druh Australopithecus afarensis. Pozn. překl.).
Naše argumenty proti těmto teoriím o bipedii najdete v předešlém výkladu v oddílu o kadabbovi.
Australopithecus anamensis
Roku 1995 ohlásili příslušní paleoantropologové další nález, tentokrát zejména vzorků zubů, lebečních a týlních kostí; pocházely ze dvou samostatných míst v Keni a jejich stáří činilo asi 3.8 až 4.2 milionu let. Vědci se domnívali, že patří novému druhu hominida pojmenovanému Australopithecus anamensis (38). Většina zkamenělých úlomků pocházela nepochybně z lidoopa, třeba šimpanzoidní čelisti, spory se však rozhořely kolem údajné antropoidní povahy kostí holeně a kostí pažních (39). Pažní kosti anamensise schází hluboký oválný otvor sloužící ke spojení s kostí loketní vyskytující se u šimpanzů, nikoli však u lidí; holenní kost anamensise je pak široká jako u lidí vzhledem k přítomnosti zvláštní houbovité tkáně tlumící otřesy při chůzi o dvou (40).
V článku s popisem anamensise se paleontolog Peter Andrews zmínil o možnosti, že jak holenní kost, tak kost pažní, pocházející z jiných profilů (svrchní etáže) vrstev lokality Kanapoi než „primitivní“ čelisti a zuby (spodní etáž), by mohly „patřit předkům člověka a zmíněný primitivní zbytek lidoopům“ (41). Ve vrstvách svrchní etáže se sice také našlo několik úlomků dolní čelisti (KNM-KP 29287), ty však pocházely z jiné (vyšší) etáže než kost holenní (KNM-KP 29285) i kost pažní (KNM-KP 271) (42).
V oné době byl také problémem paleontologů fakt, že mezi zmíněnou horní a dolní etáží leželo několik set tisíc let evolučního času; v pozdějším článku na naše téma však paleontologové přišli s daty, která tuto časovou prodlevu zmenšila (43). Tato pozdější zpráva se též zmiňovala o dalších nálezech fosilních zbytků anamensise, její autoři v ní však neuvedli žádné podrobnější informace o vztahu těchto nových nálezů ke zmíněné kosti holenní a pažní. Také analýza zkamenělé kosti vřetenní (KNM-ER 20419) z dalšího naleziště anamensise, Aliia Bay, provedená roku 2000, ukazovala na „specializovanou morfologii zápěstí svědčící o chůzi na kloubech končetin“ (44). V článku, jehož spoluautorkou byla Meave Leakeyová, se konstatovalo, že anamensis se velmi podobá afarensisovi; jeho kost týlní, pažní, holenní i vřetenní „jsou téměř stejné co do velikosti i morfologie jako kosti A. afarensis nasbírané v Hadaru“ (45).
Jestli tedy zbytky kosti týlní skutečně patří anamensisovi, pak máme s největší pravděpodobností co dělat s tvorem podobným lidoopům s podobnou lokomocí jako u pitekoidního afarensise. Pro další podrobnosti odkazujeme čtenáře na následující oddíl o afarensisovi, u něhož byla již provedena hloubková analýza nálezů zahrnujících také větší množství fosilií kosti týlní než tomu bylo u anamensise.
(Hadar = naleziště fosilních australopiteků (1972-77) na stř. toku řeky Awaš v údolí Afar v severových. Etiopii. Datování: 3.6 až 2.2 milionu let. Pozn. překl.)
Australopithecus afarensis
O nálezu kostry slavné Lucy v Etiopii roku 1974 (46) a o sporu kolem názvu Australopithecus afarensis roku 1978 (47) se psalo dostatečně a nebudeme zde tyto historky znovu vyprávět. Odhady objemu mozkové dutiny afarensise, jehož evoluční stáří sahá od 3.0 do 3.9 milionů let, se pohybují od 375 do 540 cm3 s průměrem zhruba 470 cm3 (48), ačkoli průměrná velikost mozku je v jiných pracích udávána 438 cm3 (49). Lebka afarensise nad krkem (včetně čelistí) byla popsána jako podobná lidoopům (50); evolucionističtí znalci však zároveň mluví o její odlišnosti od lebek šimpanzích hlavně s ohledem na drobnější špičáky a mohutnější řezáky afarensise „a vliv, který mají zmíněné menší špičáky na tvář tohoto tvora včetně menšího čenichu a výskytu jamek ve zmíněných špičácích „ (51).
Podrobnosti o první lebce afarensise (AL 444-2) byly zveřejněny roku 1994 (52) a v oné době to byla největší známá lebka australopiteka (53) s odhadovaným objemem mozkové dutiny stanoveným později na 540 cm3 (50). Předtím vycházela rekonstrukce lebky afarensise z fosilních zlomků několika individuí skládaných dohromady (54) a v důsledku toho obviňovali někteří vědci autory této skládanky Kimbela a Whita, že „neorganicky přimontovali tvář štíhlého hominoidního druhu na mozkovnu tvora robustního“ (55).
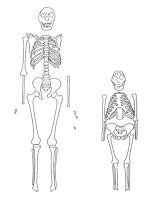
Obrázek: Srovnání kostry druhu Homo erectus KNM-WT 15000 (vlevo) s kostrou druhu Australopithecus afarensis Lucy (vpravo). Všimněte si obrovského morfologického rozdílu mezi v podstatě modernímu člověku odpovídající postkraniální kostrou (kost týlní atd.) erectuse a postkraniální kostrou australopiteka reprezentovanou Lucy. Například kuželovitý hrudní koš Lucy svědčí o ukládání tuku v oblasti pasu s vystouplým břichem podobným šimpanzímu. Také měla Lucy široce nálevkovitě rozšířené kyčelní kosti pánve ve srovnání s užšími kyčlemi lidskými. Další podrobnosti uvádí Mehlert (113). (Obrázek z publikace Walker a Shipmanová – 114).
Pokud jde o zbytek těla, vykazoval afarensis pitekoidní znaky jako třeba kuželovitý hrudní koš a spolu s dalšími australopiteky byl robustní a „téměř jistě nebyl přizpůsoben chůzi o dvou či běhu jako lidé“ (56). Lidé mají hrudní koše soudkovité a ve srovnání s lidoopy jsou subtilnější postavy. Z analýzy jamky na lopatce (scapule) afarensise vyplynulo, že lopatka směřuje daleko více vzhůru (kraniálněji) (57) ve srovnání s lidmi, jejichž jamka na lopatce směřuje naopak kolmo k zemi, a že podobně jako u lidoopů měla tato orientace jamky afarensise „smysl tehdy, měli-li tito australopitekové po většinu času paže vztyčené nad hlavu jako je tomu tehdy, když opice šplhá po stromech a zavěšuje se na ně“ (58). Pokud jde o proporce končetin, pak je odhadovaný humerofemorální index (poměr délky pažní kosti k délce kosti stehenní) u afarensise menší než obdobný index žijících lidoopů, i když stále podstatně větší než zmíněný index u lidí (59). Lidé mají relativně krátké paže a velmi dlouhé nohy a podle Tattersalla a Schwartze je naopak „jisté, že nohy afarensise byly celkem krátké“ (60). Odhadovaný brachiální index (poměr délky kosti vřetenní k délce kosti pažní) afarensise je větší než obdobný index lidí a goril, avšak menší než index ostatních žijících lidoopů; zmíněný odhad je však velmi nespolehlivý vzhledem k obtížím spojeným se snahou odhadnout přesně délku vřetenní kosti Lucy (AL 288-1) – velkorysost při zmíněném odhadu by totiž samozřejmě posunula brachiální index afarensise do kategorie šimpanzů (61). Tattersall a Schwartz však konstatují, že
„i když jsou relativní proporce paží spíše antropoidnější než pitekoidní (předloktí je ve srovnání s horní částí paže šimpanzů a zejména výhradně stromových lidoopů – orangutanů a gibbonů – mnohem delší) – byly stejně afarensisovy paže celkem dlouhé“ (60).
Afarensisovy kůstky prstů rukou, zejména jejich proximální části, jsou zakřivené, podobné šimpanzím, a „svědčí o adaptaci na zavěšování se na větve a šplhání po stromech, které předpokládá schopnost pevného úchopu“ (62). Afarensisovy prsty u nohou jsou také zakřivené (63) jako prsty lidoopů a svědčí o schopnosti šplhat po stromech. Podle Sterna a Susmanové „svědčí nalezené zbytky chodidel i kotníků afarensise jak o tom, že šplhal, tak o tom, že chodil o dvou“ (64). Oba vědci pak pokračují tvrzením, že „Neexistují důkazy o tom, že by měl kterýkoli žijící primát dlouhé, ohnuté, svalnaté dlaně i chodidla k jinému účelu než k usnadnění pobytu na stromech, ať už stálému či jen občasnému“ (64).
Analýza zkamenělin afarensisových dlaní provedená Sternem a Susmanovou roku 1983 prokázala mnoho znaků společných se šimpanzi; zmínění vědci však zároveň upozornili na fakt, že zkoumání záprstních kůstek nepřineslo důkazy o tom, že by se afarensis při chůzi opíral o klouby (65). Pozdější výzkum však zmíněné důkazy přesto přinesl. Morfologie zápěstí žijících afrických lidoopů chodících po kloubech (šimpanzů a goril) jim umožňuje sevřít při chůzi dlaň tak, aby se zabránilo „natažení zápěstí, jakmile na ně dopadne váha těla“ (44). Roku 2000 napsali Richmond a Strait, že morfologie distální části kosti vřetenní (včetně zářezu na ulně) se u Australopithecus anamensis i A. afarensis podobá téže morfologii afrických lidoopů chodících po kloubech, zatímco kost vřetenní připisovaná A. robustus a zejména A. africanus je antropoidnější (66). Oba vědci ovšem v závěru své práce překvapivě konstatovali, že zmíněné znaky vřetenních kostí anamensise i afarensise byly nefunkčními přítěžemi (zpomalujícími evoluci) zděděnými po předcích (67). Takové znaky však nejsou něčím, co by mělo u organizmu dlouho přetrvávat, podle evolucionisty Jeremyho Cherfase:
„Celá její kostra od konečků prstů rukou po prsty na nohou svědčí o tom, že u Lucy i jejích sester přetrvávaly některé znaky, které by se velmi hodily při lezení po stromech. Některé ze zmíněných znaků můžeme pak rovněž pozorovat, i když ve velmi omezené míře, i u mnohem mladších hominidů jako třeba u 2 miliony let starých exemplářů Homo habilis z olduvajské rokle. Můžete namítnout, že schopnost Lucy šplhat po stromech jí prostě zůstala po předcích, kteří žili výhradně na nich, ale nestává se přece často, že by u zvířat přetrvávaly schopnosti, jichž nevyužívají, a nález předkovských znaků u jejich potomků mladších o celé 2 miliony let nesvědčí zrovna o tom, že by šlo o pouhé atavizmy.“ (68)
(Olduvai = údolí v serengetské stepi v Tanzanii, v jehož geol. vrstvách byly nalezeny fosilní kostry antropogenetické řady. Výzkumy zde prováděl především L.S.B.Leakey a jeho žena. Stáří nálezů 30 tis. až 1.9 mil. let. Pozn. překl.)
Další zajímavou skutečností v Richmondově a Straitově studii je zjištění, že morfologie africanova zápěstí je „modernější“ než zápěstí afarensisovo. Jiné studie však zase naopak konstatují, že proporce africanova chodidla, holeně i celých končetin jsou pitekoidnější než příslušné partie u afarensise, i když paradoxně je opět jeho lebka šimpanzoidnější než lebka africanova (69). Člověk je právem na rozpacích, jaké evoluční vztahy mezi oběma druhy vlastně panovaly, vyplývají-li z poznatků vědců o jejich anatomii tak rozporné závěry; převažujícím názorem však zůstává to, že afarensis byl předkem africana. Jak píší Collard a Aiello:
„Už nejde jen o to, že by lebka svědčila o jedné škále fylogenetických vztahů, a postkraniální kostra – tedy vše kromě lebky – o jiných fylogenetických vztazích. Dokonce už i různé části postkrania svědčí o odlišných fylogenezích zmíněných dvou druhů Australopitheků.“ (69).
Pokud jde o Lucy, píše o ní profesor archeologie Clive Gamble:
„Její anatomie svědčí jednoznačně o bipedii, leč přece jen trochu odlišné od moderních lidí. Však jsou také některé anatomické rysy Lucy tak odlišné od lidí, že Charles Oxnard ve své obsáhlé studii o jejích končetinách musel konstatovat, že způsob její lokomoce nepřipomínal ani lidoopy ani lidi. A nešlo ani o jakési přechodné stadium mezi lokomocí obou zmíněných hlavních skupin; chůze Lucy byla prostě unikátním fenoménem.“ (70).

Obrázek: Lucy je nejslavnějším exemplářem druhu Australopithecus afarensis, z něhož se zachovalo zhruba 40% postkraniálních kostí.
Ústředním argumentem pro obhajobu hominidního statutu afarensise je postulát, že tento tvor měl vzpřímené držení těla a chodil po dvou. Důkazy jeho bipedie jsou založeny na rekonstrukcích pánevních kostí a kostí dolních končetin. Jelikož se našlo skoro 40% postkraniální kostry Lucy, provedli vědci na základě tohoto materiálu spoustu morfologických a biomechanických analýz, ačkoli se samozřejmě ve svých závěrech neopírají úplně výhradně jen o ně. Problémem při rekonstrukcích Lucyiny kostry (staré zhruba 3.2 milionu let) je určitý subjektivizmus, protože apriorní víra ve vzpřímené držení těla afarensisů zřejmě skutečně nepříznivě ovlivňuje její přesnou rekonstrukci. Jak jinak bychom si měli vyložit následující výroky Maurice Abitbola, který studoval Lucyinu pánev?
„Je téměř nemožné najít kompromis mezi současnými hlavními výklady otázky Lucyina držení těla. Kdybychom dali na rekonstrukci provedenou Lovejoyem (1998) a Weaverem et al. (1985), vyšla by nám postava zcela moderního bipedního člověka (obrázek 1a). Avšak podíváme-li se na její předběžně rekonstruovanou figuru zařazenou nedávno do expozice ve Smithsonianu, získáme dojem šimpanze neobratně se pokoušejícího udržet se na zadních a stále trochu padajícího na přední (Lewin, 1988). V tomto případě se tedy vnucuje závěr, že v případě Lucy šlo o „primitivní“ formu bipedie u hominidů z Hadaru. Abychom tyhle rozpory srovnali, potřebujeme více anatomického materiálu, tedy více fosilií. Současný stav výzkumů na tomto poli totiž stále nedovoluje celý problém jednoznačně uzavřít.“ (71).
(Smithsonian, Smithsonův institut = badatelská organizace ve Washingtonu, D.C. založená 1846 z odkazu anglického chemika Jamese Smithsona (1765-1829) a spravující četná muzea a galerie. Pozn. překl.)
K velkému úžasu a zmatku vědeckého světa lze tedy Lucyino držení těla interpretovat podle celé škály vzorů od moderního člověka až k šimpanzům. Kreacionisté mají tedy plné právo být krajně skeptičtí k názorům, že Lucy i jiní australopitekové byli evolučními mezičlánky mezi lidopy a lidmi, jsou-li hlavní důkazy, o něž se takové názory opírají, založeny na tak spekulativních interpretacích dat. Abitbol uzavřel svou studii konstatováním, že „Lucyino vzpřímené držení těla se lišilo od chůze moderních lidí a je stále obestřeno tajemstvím.“ (72).
Dalším problémem při výzkumu pohybových schopností fosilních tvorů jako je afarensis je fakt, že diskuze o svalových funkcích „předpokládají a priori, že šlo o svaly téměř lidské.“ (73). Po biomechanické analýze Lucyiny pánve a kostí dolních končetin (a zejména po rekonstrukci svalů gluteálních – hýžďových) napsala ovšem Christine Bergerová, že „pitekoidní hýžďové svalstvo by umožňovalo Lucy snadnější pohyb než svalstvo antropoidní“ (73). Svá studia uzavřela Bergerová takto:
„Bipedie australopiteků se musela lišit od bipedie lidské. Nejenže měl Australopithecus potíže udržet při chůzi napjaté svalstvo kyčlí a kolen, nýbrž i jeho svalstvo pánevní a svaly dolních končetin se patrně pohybovaly odlišně. Zdá se tedy, že chůze australopiteků se podstatně lišila od lidské a připomínala chůzi kolébavou se značným natáčením pánve i ramen kolem páteře (Bergeová, 1991a, b). Taková chůze ovšem zřejmě stála zmíněné hominidy více energie než bipedie lidská“ (74).
Podrobná analýza postkraniální kostry „poskytla Sternovi a Susmanové závažné důkazy o tom, že lezení po stromech a pobyt v jejich korunách byly pro A. afarensis tak důležité, že se u něho zachovaly morfologické znaky umožňující mu takové obratné pohyby“ (75). Zmínění vědci také o afarensisovi napsali, že „jeho chůze o dvou po zemi, kterou občas zkoušel, se lišila od chůze moderních lidí“ (75). Také skeny CT provedené skupinou vedenou anatomem Fredem Spoorem z liverpoolské univerzity a zaměřené na kostěný labyrint (polokruhovité kanálky s tekutinou) vnitřního ucha, který je součástí vestibulárního aparátu umožňujícího udržet rovnováhu (76), prokázaly, že rozměry zmíněných kanálků v lebkách připisovaných dalším australopitekům (africanovi a robustovi) se blížily rozměrům těchto kanálků u žijících velkých lidoopů (77). Ačkoli pro afarensise takové skeny provedeny nebyly, je pravděpodobné, že ani jeho labyrinty by se v podstatě nelišily od labyrintů šimpanzů i dalších lidoopů, jako se od nich téměř neliší labyrinty ostatních australopiteků (78). Z podobných svědectví tedy vyplývá, že afarensis téměř určitě ještě chůzi o dvou úplně nepřivykl. Také není jisté, zda nebyla omezená bipedie vyskytující se u některých australopiteků, jako třeba u afarensise, spíše pasivní a sloužící výhradně nutnému obstarávání potravy (na rozdíl od stálého aktivního pohybu, kterým se organizmus přemisťuje i na větší vzdálenosti) (79). Vzpomeňme si jen v této souvislosti na žijící bonoby (trpasličí šimpanze), u kterých byla pozorována bipední chůze po zhruba 10% jejich dne – takže bipedie se v omezené míře kromě lidí vyskytuje dokonce i u žijících lidoopů (79).
Čím je tedy vlastně přesně afarensis? Odpověď zní: tvorem s mozkem velikosti mozku lidoopího, pitekoidní lebkou a tělem připomínajícím svými rozměry i habitem rovněž lidoopa, tvorem specializovaným na lezení po stromech a chůzi o čtyřech – tvorem ve všem připomínajícím lidoopy. Jinými slovy, máme co dělat s lidoopem. Každopádně, i kdyby měl afarensis jistou omezenou schopnost pohybovat se o dvou (ovšem samozřejmě chůzí odlišnou od chůze lidské), lokomocí možná obratnější než je svrchu zmíněná chůze bonobů, je to důkazem, že šlo o opočlověka či to prostě svědčí o pestrosti habitů lidoopů/australopiteků vytvořené Bohem?
O tom, že správnou odpovědí je posledně jmenovaná možnost, svědčí analýza kosterních pozůstatků lidoopa z Itálie zvaného Oreopithecus bambolii starých od 7 do 9 milionů let, o němž snad nikdo nebude tvrdit, že šlo o hominida, jelikož zmíněné ostatky byly vykopány na špatném místě a mimo časové „okno“ hominidů – a přece jejich anatomie opravňuje k domněnce, že se s námahou „belhal“ o dvou (80). Podle vědců, kteří kosti zkoumali, se části bamboliiovy pánve podobají pánvi afarensisově a na jeho stehenní kosti se našel „zřetelný diafyzární ohyb spojený s téměř stejně velkými kondyly morfologicky podobný útvaru u rodů Australopithecus i Homo, funkčně související s bipedními aktivitami“ (81).
(stehenní kost – femur = nejdelší kost v těle tvořící podklad stehna. Shora dolů ji dělíme na tyto části: epifýzu, diafýzu a metafýzu. Kondylus = rozšířený kloubní konec kosti. Pojem se užívá u vzdálených (distálních) konců kosti pažní a stehenní, a u dolní čelisti. Každý kondylus má na vnitřní i vnější straně kostěné výstupky, na které se upínají šlachy svalů. Pozn. překl.)
Podle Henryho Gee „soudí vědecký svět, že se u tohoto tvora vyvinula bipedie samostatně a že byl pouhým vzdáleným příbuzným hominidů“ (82). Je již tak dost obtížné představit si či uvěřit tomu, že se u primátů kdysi vyvinula jistá forma bipední lokomoce byť jen jednou; ale víra, že k tomuto kroku muselo docházet samostatně mnohokrát, aby byla evoluční teorie zachráněna, svědčí o tom, že evoluce je pouhou sbírkou mýtů aplikovatelných prakticky na všechny scénáře – ať už sebeabsurdnější nebo sebenepravděpodobnější. Skutečnost, že omezená a pro lidi netypická forma bipedního pohybu existovala u lidoopů mnohem dříve a bez vztahu k nějakým hypotetickým hominidům svědčí o tom, že přítomnost bipedních rysů není rozhodujícím kritériem pro přiznání statutu hominida.
Kenyanthropus platyops
V březnu 2001 se na obálce Nature objevil další „hominid“, s titulkem hlásajícím, že se „Lidská rodina rozrůstá“. Časopis pak přinášel článek popisující nového tvora od jezera Turkana zvaného Kenyanthropus platyops (83).
(Turkana = dříve Rudolfovo jezero, bezodtokové slané jez. ve vých. Africe, v hraniční obl. Keni a jejích sev. sousedů Súdánu a Etiopie – název podle národa v sev. Keni. Rozloha 6405 km2, ve výšce 375 m n. m., až 73 m hluboké. V pliocénních a pleistocénních jezerních sedimentech (starých 5 milionů až 0.5 milionu let dle evoluční chronologie) záp. a vých. pobřežních obl. byly nalezeny četné zbytky dávných předchůdců člověka, jak věří evolucionisté. Pozn. překl.)
Se svým stářím 3.5 milionu let soupeří platyops bezprostředně s afarensisem o status hominida hlavní linie, která prý vyústila v rod Homo (84). Největší rozruch přitom vyvolala mezi vědci jeho silně deformovaná lebka. Objem její mozkové dutiny přirovnali sice k objemu této dutiny u primátů, leč zejména vzhledem k její ploché lícní straně neobvyklé u údajného hominida v tomto evolučním stadiu zařadili nalezený exemplář do nového rodu Kenyanthropus. Existují však závažné důvody k tvrzení, že ke zploštění tlamy došlo vinou prudké deformace expandující základní hmoty kostí, tedy že jde o strukturu vzniklou druhotně v procesu fosilizace (85), takže platyops je nepochybně prostě jen dalším primátem.
(expanze = růst určitého útvaru v těle, který utlačuje okolní struktury, ale neprorůstá do nich – opak invaze. Takto rostou např. benigní nádory. Pozn. překl.)
Australopithecus bahrelghazali
Roku 1995 se našla v Čadu dolní čelist tvora označeného později za nejbližšího příbuzného afarensise, který žil v téže době (86). Exemplář starý cca 3 až 3.5 milionu let zařadili vědci roku 1996 do nového druhu Australopithecus bahrelghazali. Zjistily se určité rozdíly mezi ním a afarensisem v morfologických znacích zmíněné dolní čelisti, kořenů třenových zubů i v tloušťce skloviny těchto premolárů. Vcelku však takový skrovný nález sotva opravňuje k postulování nového druhu. Není proto důvod považovat bahrelghazaliho za cokoli víc než pouhého primáta, patrně patřícího k témuž druhu jako afarensis.
Australopithecus africanus
Průměrná velikost mozku sedmi exemplářů druhu Australopithecus africanus je 451 cm3, s celkovým rozpětím od 425 cm3 do 515 cm3 (87). Evolucionisté mají za to, že africanus žil v jižní Africe před zhruba 2,5 až 3 miliony let, ačkoli je možné, že existoval až do doby před 2 miliony let (88). Pokud jde o hlavovou část těla, spatřují vědci hlavní odlišnost morfologie africana ve srovnání s afarensisem v lícních kostech – ty jsou u africana méně prognatické a širší. V porovnání s afarensisem je také africanova dolní čelist robustnější, jeho zuby třenové a stoličky jsou mohutnější a zuby přední drobnější (89).
prognatismus – dopředu vybíhající alveolární /dásňové/ výběžky horní čelisti, poz. edit.)

Obrázek: Lebka druhu Australopithecus africanus STS 5 (přezdívaná „paní Plesová“). Je jednou z nejúplněji zachovaných lebek australopiteků, které kdy byly nalezeny. Byla objevena roku 1947 Robertem Broomem a Johnem Robinsonem ve Sterkfonteinu v Jihoafrické republice. Objem její mozkové dutiny činí zhruba 485 cm3. Foto: David Green.
(Sterkfontein = naleziště rodu Australopithecus ze spodního pleistocénu. Jak věří evolucionisté, tito hominidi již používali kamenné (křemenné) nástroje nacházené spolu s jejich kostmi v již. a vých. Africe; šlo o hrubě otloukané kamenné valouny, ostřené nebo škrábané nástroje. Datování je až 2.6 milionu let. Pozn. překl.)
Mnozí badatelé považují druh Australopithecus afarensis za africanova předka, leč ze srovnání proporcí končetin obou druhů vyplývá, že údajně starší a kraniodentálně „primitivnější“ afarensis má méně pitekoidní klouby na předních končetinách než na zadních a obecně méně pitekoidní délku končetin (90). Podle zmíněných badatelů „To znamená, že proporce končetin v průběhu evoluce hominidů kolísaly k nižším a opět vyšším hodnotám či že naše současné chápání vztahů mezi hominidy je příliš zjednodušené“ (91).
Podle názoru autora tohoto článku vyplývá zmatek v chápání zmíněných vztahů spíše z faktu, že k evoluci hominidů nikdy nedošlo. Podle Kleina a Edgara byla africanova horní část těla značně pitekoidní, s dlouhými, mohutnými pažemi vhodnými pro lezení po stromech, avšak morfologie části dolní svědčila o „habituální bipední lokomoci v terénu“ (92). Jak jsme se však již zmínili, prokázaly skeny CT kostěného labyrintu vnitřního ucha fakt, že rozměry polokruhovitého kanálku v lebkách připisovaných africanovi se podobají dimenzím tohoto útvaru u žijících velkých lidoopů (77). I kdyby tedy přece jen africanus nějakým způsobem chodil po dvou, není pravděpodobné, že mu byl takový způsob pohybu bytostně vlastní.
Analýza těl afarensisů i africanů od lebky dolů (provedená v počátečních stadiích zkoumání zmíněných hominidů) vedla McHenryho k závěru,
„že (1) postkraniální kostra raných australopiteků vykazuje zcela svébytné znaky naprosto odlišné od jakéhokoli žijícího lidoopa či člověka; (2) zmíněná kostra raného A. africanus se velmi podobá kostře A. afarensis, přestože se jinak jejich lebky, uspořádání zubů a celkové stáří liší a (3) lokomoční adaptace těchto hominidů se ustálila na zcela specifické úrovni, kterou samozřejmě nepozorujeme u žádného žijícího tvora“ (93).
Jestliže se tedy u obou zmíněných druhů hominidů vyskytovala velmi podobná „jedinečná konfigurace, ze které vyplývá, že jejich lokomoční i posturální /pro přímé držení těla/ adaptace probíhala téměř shodným zcela specifickým způsobem, který dnes samozřejmě u žádného žijícího tvora nepozorujeme a pozorovat ani nemůžeme“ (94), pak se lokomoční chování africana podobalo afarensisovu.
Jak jsme shora konstatovali, byl podle našeho názoru afarensis lidoopem užívajícím kotníkovou chůzi a stavěným pro život na stromech, ačkoli možná měl omezenou schopnost chůze po dvou (ovšem samozřejmě způsobem zcela odlišným od dnešního člověka) – tak jak to platí i o dalších lidoopech, kteří nemají co dělat s údajnou evolucí hominidů, jako je třeba Oreopithecus bambolii. Stejně jako všichni ostatní australopitekové měl podle našeho názoru i africanus mozek velký jako dnešní lidoopi, měl pitekoidní lebku a svou morfologií i celkovou velikostí těla se také podobal dnešním lidoopům. Nejpřijatelnějším vysvětlením tedy zůstává, že africanus byl druhem vymřelého lidoopa; dokonce i mezi evolucionisty se diskutovalo o tom, že africanus a afarensis „by měli být považováni za poddruhy jediného druhu“ (95).
Australopithecus garhi
Roku 1999 prohlásili vědci ostatky druhu Australopithecus garhi, údajně 2.5 milionu let starého hominida z vrstev Hata na středním toku řeky Awaš v Etiopii, za odlišné od afarensise „svými daleko mohutnějšími třenovými zuby a stoličkami i morfologií jednoho z třenových zubů horní čelisti s redukovaným meziobukálním spojením v čáře skloviny a méně asymetrickým skusem“ (96).
(meziální = bližší středu zubního oblouku. Užívá se zejm. pro označení plošek zubu nebo stěn zubní dutiny. Opak: distální. Bukální = týkající se tváře a její sliznice, tvářový. Jako bukální se někdy označují plošky zubů směřující ke rtům. Pozn. překl.)
Vezmeme-li však v úvahu fakt, že ze zmíněného tvora se zachovalo vlastně jen velmi málo kraniálních zbytků a že se objem jeho mozkové dutiny odhaduje na 450 cm3, je těžké ubránit se pochybnostem o tom, zda příslušní vědci nekladli až příliš velký důraz na rozdíly v několika málo znacích zubů – vždyť by bylo jistě možné zařadit tenhle exemplář ke druhu afarensis. Snad zde rozhodovalo to, že garhi představuje v evoluci hominidů přece jen poněkud mladší fázi. Bez ohledu na jeho příbuznost s afarensisem svědčí stejně pitekoidní morfologie lebky garhiho spíše o jeho příslušnosti k vývojové linii lidoopů než člověka.
Zajímavější však byly postkraniální zkameněliny (BOU-VP-12/1) vykopané na shora zmíněném místě, které badatelé nepřipsali garhimu, protože jejich humerofemorální index svědčí tentokrát spíše o příslušnosti k vývojové linii člověka (97). Odhadovaný brachiální index kostry Bouriho (BOU-VP-12/1) je pak rovněž vyšší než tentýž index všech žijících lidoopů s výjimkou orangutanů (Pongo pygmaeus) (98). Tahle kombinace dlouhého pitekoidního předloktí s antropoidními humerofemorálními proporcemi končetin tedy přece jen v tomto případě poněkud ztěžuje stanovení přesných evolučních vztahů předek-potomek, jelikož „se zdá, že svědčí o eventuálním návratu Bouriho (přesněji proporcí jeho paží) do vývojové linie opic, tedy před A. afarensis, přičemž zase po Bourim by podle ostatních znaků jeho kostry následoval další zvrat v H. ergaster (více lidský)“ (99).
Robustní australopitekové
Mezi robustní australopiteky řadíme obvykle druhy Australopithecus robustus ( starý 1.5 až 1,8 milionu let) z jižní Afriky a dva druhy východoafrické, Australopithecus boisei (starý 1,3 až 2,3 milionu let) a Australopithecus aethiopicus (starý 2,3 až 2,7 milionu let) – celkový evoluční časový záběr zmíněných tří druhů sahá tedy od 1,3 po 2,7 milionu let (100). Někteří paleoantropologové pro ně dokonce vymysleli i zvláštní rod, Paranthropus. Žádný z robustních australopiteků není podle evolucionistů předkem pravých lidí; tito tvorové byli spíše postranní větví, jejíž vývoj se zastavil (101), protože „byly jejich zuby i lebky příliš specializované a protože je před 2,5 miliony let překonali a vytlačili pružnější, všestrannější a méně specializovaní hominidi“ (102). Jejich pojmenování „robustní australopitekové“ staví na faktu, že měli „mohutné zuby, silnou žvýkací svalovinu a masivní lebky“; jinak byly ovšem jejich mozky i těla malých rozměrů (103). Lebky těchto tvorů svědčí o tom, že na nich nic lidského nebylo.
Australopithecus aethiopicus
Slavná „černá lebka“ (KNM-WT 17000) s objemem mozkové dutiny zhruba 410 cm3 je jednou ze tří či čtyř vykopaných kostí, které tvoří ostatky druhu aethiopicus (104) považovaného některými evolucionisty za vhodného kandidáta na spojovací článek mezi afarensisem a dvěma dalšími robustními druhy australopiteků, boiseiem a robustem (105). Zmíněná lebka však podle některých vědců „vykazuje naopak až v extrémní míře znak A. robusta/boiseie : masivní sagitální hřeben; oba nalezené zuby jsou pak čtyřikrát nebo pětkrát větší než lidské stoličky“ (106), a tak i když je aethiopicus údajně vývojově starší než robustus/boisei, činí ho uvedená hyperrobustnost nezpůsobilým pro status přechodné formy. Svou morfologií se „černá lebka“ v mnohém podobá lebce malého gorilího samce (107), takže není vyloučeno, že gorily i aethiopicus patří k témuž biblickému druhu. V současné době neexistují žádné postkraniální zkameněliny aethiopicuse – jeho zbývající dvě (možná tři) objevené kosti představují totiž dolní čelisti (108).
Australopithecus boisei
Průměrná velikost mozku tohoto robustního australopiteka (soudě podle šesti zachovaných exemplářů) činí zhruba 452 cm3, s rozpětím od 390 cm3 do 500 cm3 (109); jiní vědci však odhadují zmíněnou střední hodnotu až na 521 cm3 (49). Pokud jde o lebku jako celek, popisují vědci boiseie jako „jediného hominina, u kterého se spojuje masivní, široká, plochá tvář s nevelkým neurokraniem“ (110). Neexistují sice žádné postkraniální ostatky, které lze s určitostí připsat boiseiovi, v lokalitě Koobi Fora byly však vykopány části kostry a některé postkraniální kosti, o kterých vědci přece jen uvažují jako o eventuálních zbytcích zmíněného druhu (110).
(Koobi Fora = paleolitická lokalita v Keni severovýchodně od jezera Turkana, kde byly nalezeny kosterní pozůstatky australopiteků. Datování: 2 až 3 miliony let. Pozn. překl.)
Podle Wooda a Richmonda „charakterizují zmíněné části kostry proporce končetin připomínající A. afarensis (Grausz et al., 1988) a jsou méně pitekoidní než tytéž proporce u A. africanus“ (110). Patřila-li tedy ona neúplná kostra (KNM-ER 1500) opravdu boiseiovi, pak se zřejmě pohyboval podobně jako afarensis a „podobně jako u A. afarensis svědčí i jeho relativně dlouhé přední končetiny o tom, že jeho lokomoční chování zahrnovalo arborikolní složku“ (111). /arborikolní – mající vztah ke stromům/
Australopithecus robustus
Objem mozku tohoto tvora, který zatím můžeme odhadovat pouze z jediného exempláře (SK 1585), činil 530 cm3; novější odhad však tuto hodnotu snížil na 476 cm3 (109). Jinak připomínala lebka robusta lebku boiseie, její lícní strana však nebyla tak velká a široká (110). Označíme-li tedy lebku robusta za „robustní“, pak byla lebka boiseie opravdu hyperrobustní. Ve srovnání s africanem byla robustova žvýkací svalovina i tvář mohutnější, byť byly jeho špičáky a řezáky menší (112) a podle Kleina a Edgara měly oba druhy
„zřetelně pitekoidní horní polovinu trupu s dlouhými, silnými pažemi, které svědčily o jejich potenciální hbitosti při šplhání po stromech. Od dryopiteků (vymřelých lidoopů) se pak lišili zejména dolní polovinou trupu přizpůsobenou habituální bipední lokomoci po zemi a svými zuby“ (92).

Foto: Warwick Armstrong
Obrázek: Jak je patrné z této rekonstrukce lebky Australopithecus boisei, nepřipomínaly hlavy robustních australopiteků v ničem hlavu člověka. Do podobného sádrového odlitku téže lebky v Muzeu člověka v San Diegu zapracovali vědci i dolní čelist, jejíž tvar odvodili z fosilie nalezené o cca 75 km dále.
Podle Wooda a Richmonda připomíná morfologie pánve a kyčle (o které se soudilo, že bude bližší robustovi ) podobné struktury afarensise a africana („gracilních“ australopiteků) a svědčí o tom, že „chůze druhu P. robustus se patrně podobala chůzi ´gracilních´ australopiteků“ (112). Jak jsme se zmínili dříve, lišila se afarensisova chůze od chůze lidské. A jak jsme také již dříve zdůraznili, prokázaly CT skeny kostěného labyrintu vnitřního ucha, že rozměry polokruhovitého kanálku v lebkách připisovaných robustovi připomínaly podobné parametry žijících velkých lidoopů (77).
Závěr
Morfologické znaky hominidních fosilií připisovaných taxonům neřazeným k rodu Homo svědčí o tom, že byly pouhými vymřelými lidoopy. Současný biologický druh /species/ není totožný s druhem biblickým, a tak snad představovali někteří ze zmíněných hominidů prostě varianty v rámci jednoho či více lidoopích druhů, právě tak, jako fosilní druhy zahrnované do rodu Homo (kromě neplatného taxonu Homo habilis) představují varianty jediného druhu člověk. Lze si docela dobře představit, že žijící lidoopi jsou přežívajícími zbytky zmíněných druhů hominidních lidoopů, kteří ztratili značnou část své genetické diverzity.
Obecně můžeme konstatovat, že tam, kde máme k dispozici fosilní materiál, zjišťujeme z něho, že mozky těchto hominidních tvorů odpovídaly svým objemem právě mozkům dnešních lidoopů, morfologie jejich lebek byla pitekoidní /opičí/ a jejich trupy se podobaly lidoopím svými tvary i výškou. Morfologie tvorů jako byl Australopithecus afarensis svědčí o tom, že byli specializováni na šplhání po stromech i na chůzi po kloubech všech čtyř končetin podobně jako dnešní lidoopi. Zároveň se trochu přeceňuje význam těch znaků koster, které zdá se nasvědčují tomu, že někteří australopitekové včetně afarensise byli také v omezené míře schopni bipední lokomoce (ovšem odlišné od té lidské). Podobná omezená bipedie se však rovněž vyskytovala u vymřelých lidoopů neřazených k hominidům, jako byl třeba Oreopithecus bambolii, datovaný do doby před údajným rozštěpením vývojových linií člověka a šimpanze. Jak můžeme potom nějaký takový rys používat jako důkaz pro jedinečnost všech hominidů i jejich evoluční příbuznost, není-li zmíněný rys pro ony údajné hominidy vůbec specifický? To je jako bychom tvrdili, že se velryby vyvinuly z tvorů podobných hrochům, protože oba umějí plavat (nebo jsou snad evolucionisté přesvědčeni i o tomhle?). Mám za to, že právě z tohoto důvodu mají evolucionisté tak rádi slovo paralela a paralelní evoluce. Nazvěte to, jak chcete, ale je tohle věda?
O autorovi: Peter Line promoval z biofyziky. Poté obhájil magisterský a Ph.D. titul, obojí v oblasti neurovědy. Od doby, kdy se stal křesťanem, měl stálý zájem o otázky kreace/evoluce, protože předtím byla evoluční teorie pro něj překážkou víry v pravdivost Božího slova.
Odkazy na literaturu
1. Harrub, B. and Thompson, B., The Truth About Human Origins, Apologetics Press Inc., Montgomery, AL, pp. 8–10, 2003.
2. Gribbin, J. and Cherfas, J., The First Chimpanzee: In Search of Human Origins, Penguin Books, London, pp. 116–117, 2001.
3. Sarfati, J., Refuting Compromise, Master Books, Green Forest, AR, pp. 376–388, 2004; Lubenow, M.L., Bones of Contention: A Creationist Assessment of Human Fossils, Revised and Updated, Baker Books, Grand Rapids, MI, pp. 273–292, 2004.
4. Harrub and Thompson, ref. 1, pp. 99–134; Lubenow, ref. 3, pp. 222–235. Lubenow, ref. 3, p. 187.
5. Brunet, M. et al., A new hominid from the upper Miocene of Chad, Central Africa, Nature 418:145–151, 2002.
6. Wood, B., Hominid revelations from Chad, Nature 418:134, 2002.
7. BBC News, Skull find sparks controversy, , 13 July 2002.
8. Montgomery, B., Man’s ancestors 7m years old, The Australian, 11 July 2002, p. 1.
9. Wolpoff, M.H., Senut, B., Pickford, M. and Hawks, J., Sahelanthropus or ‘Sahelpithecus’? Nature 419:581–582, 2002.
10. Ancient skull more ape than human? , 10 October 2002.
11. Wolpoff et al., ref. 10, p. 582.
12. Brunet et al., ref. 6, p. 151.
13. Murdock, M., Sahelanthropus tchadensis—the ambiguous ape, TJ 18(3):114, 2004.
14. Beauvilain, A. and Guellec, Y.L., Further details concerning fossils attributed to Sahelanthropus tchadensis (Toumai), South African J. Science 100:142–144, 2004.
15. Beauvilain and Guellec, ref. 15, p. 142.
16. Dalton, R., Brickbats for fossil hunter who claims skull has false tooth, Nature 430:956, 2004.
17. Aiello, L.C. and Collard, M., Our newest oldest ancestor? Nature 410:526–527, 2001.
18. Balter, M., Scientists spar over claims of earliest ancestor, Science 291:1460, 2001.
19. Wong, K., An ancestor to call our own, Scientific American Special 13(2):9, 2003.
20. Begun, D.R., The earliest hominins—is less more? Science 303:1479, 2004. Shipman, P., Hunting the first hominid, American Scientist 90(1), 2002; , 13 August 2004.
21. Galik, K., Senut, B., Pickford, M., Gommery, D., Treil, J., Kuperavage, A.J. and Eckhardt, R.B., External and internal morphology of the BAR 1002’00 Orrorin tugenensis femur, Science 305:1450–1453, 2004. Gibbons, A., Oldest human femur wades into controversy, Science 305:1885, 2004.
22. Halle-Selassie, Y., Late Miocene hominids from the Middle Awash, Ethiopia, Nature 412:178–181, 2001.
23. Halle-Selassie, Y., Suwa, G. and White, T.D., Late Miocene teeth from Middle Awash, Ethiopia, and early hominid dental evolution, Science 303:1503–1505, 2004.
24. Halle-Selassie, ref. 25, p. 180.
25. Wong, ref. 20, pp. 8–9.
26. Begun, ref. 21, pp. 1478–1480.
27. Lemonick, M.D. and Dorfman, A., One giant step for mankind, Time (South Pacific) pp. 58–59, 23 July 2001.
28. Balter, M. and Gibbons, A., Another emissary from the dawn of humanity, Science 293:189, 2001.
29. White, T.D., Suwa, G. and Asfaw, B., Australopithecus ramidus, a new species of early hominid from Aramis, Ethiopia, Nature 371:306–312, 1994.
30. White et al., ref. 32, p. 88.
31. White et al., ref. 32, p. 311.
32. White et al., ref. 32, p. 312.
33. Fischman, J., Putting our oldest ancestors in their proper place, Science 265:2011, 1994.
34. Semaw, S. et al., Early Pliocene hominids from Gona, Ethiopia, Nature 433:304, 2005.
35. Leakey, M.G., Feibel, C.S., McDougall, I. and Walker, A., New four-million-year-old hominid species from Kanapoi and Allia Bay, Kenya, Nature 376:565–571, 1995.
36. Leakey, M. and Walker, A., Early hominid fossils from Africa, Scientific American 276(6):63–64, 1997.
37. Leakey and Walker, ref. 39, p. 63.
38. Andrews, P., Ecological apes and ancestors, Nature 376:556, 1995.
39. Leakey, M.G., Feibel, C.S., McDougall, I., Ward, C. and Walker, A., New specimens and confirmation of an early age for Australopithecus anamensis, Nature 393:62, 1998; The distal end of the humerus was discovered in 1965 and is described in: Patterson, B. and Howells, W.W., Hominid humeral fragment from early Pleistocene of Northwest Kenya, Science 156:64–66, 1967.
40. Leakey et al., ref. 42, p. 62.
41. Richmond, B.G. and Strait, D.S., Evidence that humans evolved from a knuckle-walking ancestor, Nature 404:382, 2000.
42. Ward, C., Leakey, M. and Walker, A., The new hominid species Australopithecus anamensis, Evolutionary Anthropology 9:201, 1999.
43. Reader, J., Missing Links: The Hunt for Earliest Man, Penguin Books, London, pp. 207–210, 1988.
44. Lewin, R., Bones of Contention, Penguin Books, London, pp. 268–300, 1987.
45. Wood, B. and Richmond, B.G., Human evolution : taxonomy and paleobiology, J. Anatomy 196:29, 2000.
46. McHenry, H.M. and Coffing, K., Australopithecus to Homo: transformations in body and mind, Annual Review of Anthropology 29:127, 2000.
47. Tattersall, I. and Schwartz, J.H., Extinct Humans, Westview Press, New York, p. 88, 2001.
48. Wood and Richmond, ref. 48, p. 28.
49. Kimbel, W.H., Johanson, D.C. and Rak, Y., The first skull and other new discoveries of Australopithecus afarensis at Hadar, Ethiopia, Nature 368:449–451, 1994.
50. Aiello, L.C., Variable but singular, Nature 368:399, 1994.
51. Kimbel, W.H., White, T.D. and Johanson, D.C., Cranial morphology of Australopithecus afarensis : a comparative study based on a composite reconstruction of the adult skull, American J. Physical Anthropology 64:337–388, 1984; Kimbel, W.H. and White, T.D., A revised reconstruction of the adult skull of Australopithecus afarensis, J. Human Evolution 17:545–550, 1988.
52. Shreeve, J., ‘Lucy,’ crucial early human ancestor, finally gets a head, Science 264:34, 1994.
53. Leakey, R. and Lewin, R., Origins Reconsidered: In Search of What Makes us Human, Abacus, London, pp. 193–196, 1992.
54. Stern, J.T. and Susman, R.L., The locomotor anatomy of Australopithecus afarensis, American J. Physical Anthropology 60:284–285, 1983.
55. Cherfas, J., Trees have made man upright, New Scientist 97:172, 1983.
56. Richmond, B.G., Aiello, L.C. and Wood, B.A., Early hominin limb proportions, J. Human Evolution 43:534–535, 2002.
57. Tattersall and Schwartz, ref. 50, p. 90.
58. Richmond et al., ref. 59, pp. 542–543.
59. Stern and Susman, ref. 57, p. 284.
60. Stern and Susman, ref. 57, p. 307.
61. Stern and Susman, ref. 57, p. 308.
62. Stern and Susman, ref. 57, pp. 283–284.
63. Richmond and Strait, ref. 44, p. 383.
64. Richmond and Strait, ref. 44, pp. 383–384.
65. Cherfas, ref. 58, p. 176.
66. Collard, M. and Aiello, L.C., From forelimbs to two legs, Nature 404:339, 2000.
67. Gamble, C., Timewalkers: The Prehistory of Global Colonization, Penguin Books, London, p. 58, 1993.
68. Abitbol, M.M., Lateral view of Australopithecus afarensis: primitive aspects of bipedal positional behavior in the earliest hominids, J. Human Evolution 28:228, 1995.
69. Abitbol, ref. 71, p. 211.
70. Berge, C., How did australopithecines walk? A biomechanical study of the hip and thigh of Australopithecus afarensis, J. Human Evolution 26:270, 1994.
71. Berge, ref. 73, p. 271.
72. Stern and Susman, ref. 57, p. 313.
73. Shipman, P., Those ears were made for walking, New Scientist 143:27, 1994.
74. Spoor, F., Wood, B. and Zonneveld, F., Implications of early hominid labyrinthine morphology for evolution of human bipedal locomotion, Nature 369:645, 1994.
75. Gibbons, A., Anthropologists take the measure of humanity, Science 264:350, 1994.
76. Lubenow, M.L., Paleoanthropology in review, TJ 10(1):13, 1996.
77. Kohler, M. and Moya-Sola, S., Ape-like or hominid-like? The positional behavior of Oreopithecus bambolii reconsidered, Proc. Nat. Acad. Sci. USA 94:11747–11750, 1997.
78. Kohler and Moya-Sola, ref. 80, p. 11747.
79. Gee, H., Return to the planet of the apes, Nature 412:131, 2001.
80. Leakey, M.G., Spoor, F., Brown, F.H., Gathogo, P.N., Klarie, C., Leakey, L.N. and McDougall, I., New hominin genus from eastern Africa shows diverse middle Pliocene lineages, Nature 410:433–440, 2001.
81. Wong, K., Finding Homo sapiens’ lost relatives, Scientific American 285(4):26–27, 2001.
82. White, T., Early hominids—diversity or distortion? Science 299:1994–1997, 2003.
83. Brunet, M., Beauvilain, A., Coppens, Y., Heintz, E., Moutaye, A.H.E. and Pilbeam, D., The first australopithecine 2,500 kilometres west of the Rift Valley (Chad), Nature 378:273–275, 1995.
84. Falk, D., Redmond Jr, J.C., Guyer, J., Conroy, G.C., Recheis, W., Weber, G.W. and Seidler, H., Early hominid brain evolution: a new look at old endocasts, J. Human Evolution 38:709, 2000.
85. Klein, R.K. and Edgar, B., The Dawn of Human Culture, John Wiley & Sons Inc., New York, p. 35, 2002.
86. Wood and Richmond, ref. 48, p. 32.
87. McHenry, H.M. and Berger, L.R., Body proportions in Australopithecus afarensis and A. africanus and the origin of the genus Homo, J. Human Evolution 35:1–22, 1998; McHenry, H.M. and Berger, L.R., Limb lengths in Australopithecus and the origin of the genus Homo, South African J. Science, 94 :447–450, 1998.
88. McHenry and Berger, ref. 90, p. 447.
89. Klein and Edgar, ref. 88, p. 37.
90. McHenry, H.M., The first bipeds: a comparison of the A. afarensis and A. africanus postcranium and implications for the evolution of bipedalism, J. Human Evolution 15:179, 1986.
91. McHenry, ref. 93, p. 186.
92. McHenry, ref. 93, p. 187.
93. Asfaw, B., White, T., Lovejoy, O., Latimer, B., Simpson, S. and Suwa, G., Australopithecus garhi : a new species of early hominid from Ethiopia, Science 284:631, 1999.
94. Asfaw et al., ref. 96, pp. 632–633.
95. Richmond et al., ref. 59, p. 542.
96. Richmond et al., ref. 59, p. 543.
97. McCollum, M.A., The robust australopithecine face: a morphogenetic perspective, Science 284:301, 1999.
98. Johanson, D. and Edgar, B., From Lucy to Language, Simon & Schuster Editions, New York, p.152, 1996.
99. Klein and Edgar, ref. 88, p. 41.
100. Klein and Edgar, ref. 88, p. 39.
101. Johanson and Edgar, ref. 101, pp. 23, 152.
102. Klein and Edgar, ref. 88, pp. 57–58.
103. Mehlert, A.W., A review of the present status of some alleged early hominds, TJ 6(1):32, 1992.
104. Mehlert, ref. 106, p. 34.
105. Johanson and Edgar, ref. 101, p. 23.
106. Falk et al., ref. 87, pp. 708–709.
107. Wood and Richmond, ref. 48, p. 37.
108. Wood and Richmond, ref. 48, p. 38.
109. Wood and Richmond, ref. 48, p. 35.
110. Mehlert, A.W., Australopithecines—the extinct southern apes of Africa: a fresh light on their status? TJ 14(3):91–99, 2000.
111. Walker, A. and Shipman, P., The Wisdom of Bones: In Search of Human Origins, Weidenfeld and Nicolson, London, pp. 192, 1996.