
(Z creation.com přeložil .M.T. – 04/2014. Článek vyšel nejprve v časopisu CMI Journal of Creation 23(2):59-63). Translation granted by Creation.com – přeloženo s povolením od Creation.com.
Vývojové řady koní jsou již dlouho výkladní skříní evoluce. Ale ve skutečnosti tato skupina představuje nejlepší argument, který může být předložen proti evoluci z fosilního záznamu1. Kreacionisté mají různé názory na to, zda skupinu koně tvoří ve skutečnosti různé stvořené druhy. Tento článek rozebírá některé ze současných problémů a dospívá k závěru, že řada koně zahrnuje patrně tři odlišné stvořené druhy, nikoli všechna zvířata známá pod označením Hyracotherium. Zdá se totiž, že samotné Hyracotherium obsahuje několik odlišných stvořených druhů jako třeba zvířata podobná tapírům.
Fosilie koní byly nalezeny ve vrstvách sedimentů ze začátku období terciéru (třetihor), konkrétně z období zvaného eocén (přibližně před 50 miliony let podle evolucionistického datování). Obvykle jsou označovány2 jako Eohippus či Hyracotherium (viz obrázek 1).

Obrázek 1. Evoluční rodokmen koně sestavený Georgem Gaylordem Simpsonem roku 1951. Rodokmen byl později zjednodušen5, ale nedávno v něm naopak přibyly větve a je ještě chaotičtější vinou doplnění o další členy jako důsledku nových fosilních nálezů (viz odkaz č. 2). Možné evoluční mezery zde označujeme otazníkem. Equus = moderní kůň.
Podle evoluční teorie je možné sledovat vývoj koně po miliony let: jak se kůň pomalu stával větším a silnějším (obrázek 1), ztrácel mnoho svých prstů (obrázek 2) a jak se měnila struktura jeho zubů, když se přesouval od konzumování listí bylin, keřů a stromů (spásání) ke konzumování tvrdé, suché trávy (pastvě) (obrázek 3)3, 4. Má se za to, že motorem evoluce koně bylo ochlazování podnebí a přechod k suššímu podnebí. Dřívější koně žili údajně ve vlhkých lesích plných rostlin bohatých na listy. Jejich prsty, čtyři vpředu a tři vzadu, byly nepravidelně roztažené, což jim pomáhalo, aby se nepropadali v bahnitém terénu. S tím, jak se podnebí stávalo sušším, mizely rostliny s listy a tvořily se rozsáhlé travnaté savany. To nutilo pasoucí se zvířata stávat se rychlejšími běžci, aby byla schopna unikat svým predátorům.
Všichni koně jsou si navzájem natolik podobní, že byli zařazeni do jedné čeledi – koňovití. Kvůli této blízké podobnosti může být proto často obtížné rozpoznat nějaké odlišnosti pouhým studiem fosilních koster. Dalším problémem při identifikaci fosilií obratlovců naopak je, že odchylky ve strukturách dokonce i v rámci rodu žijících zvířat mohou často být tak velké, že se překrývají s odchylkami v jiných skupinách; např. mezi různými šelmami existuje mnoho analogií ve struktuře zubů, i když nejsou taková zvířata řazena do téhož rodu (nebo někdy ani do téže čeledi). Nejdůležitější diagnostické rozdíly mezi různými skupinami zvířat jsou tak často v konstrukci měkkých částí. Mnoho nálezů fosilních koní navíc tvoří pouze zuby nebo části čelistí.
Koňské skupiny
V řadě koně můžeme rozlišit určitá zvířata, která by mohla představovat stvořené druhy, i když máme k dispozici jen fosilní kostry. Zdá se, že následující fakta takovouto interpretaci podporují.
V řadě koně existují přinejmenším dvě evoluční mezery
a) První mezera se vyskytuje u Epihippa8
Z tohoto zvířete se zachovaly jen nepatrné fosilní zbytky, které se podobají zbytkům dřívějšího Orohippa, Eohippa i dalších již dříve popsaných hyracotheroidních druhů9.
b) Druhá mezera se vyskytuje u nebo hned po skupině Parahippa10
Předpokládá se, že rané druhy Parahippa se podobaly Miohippovi a Mesohippovi, zatímco o těch pozdějších se soudí, že vypadaly jako Merychippus; tyto dohady se ovšem jen zčásti opírají o fosilní nálezy11. Navíc je fosilní materiál pro Parahippa neúplný12. Bylo by patrně možné zařadit různé části Parahippa ke dvěma různým zvířatům – Miohippovi (obrázek 4) a Merychippovi13. Uvedený závěr lze rovněž opřít o práci Cavanaugha et al.14, jelikož Parahippus vykazuje podobnosti se 14 z 18 druhů koní. Proto se zdá, že stupeň „Parahippus“ v řadě koně je skupinou vytvořenou ze směsi nepříbuzných fosilií.
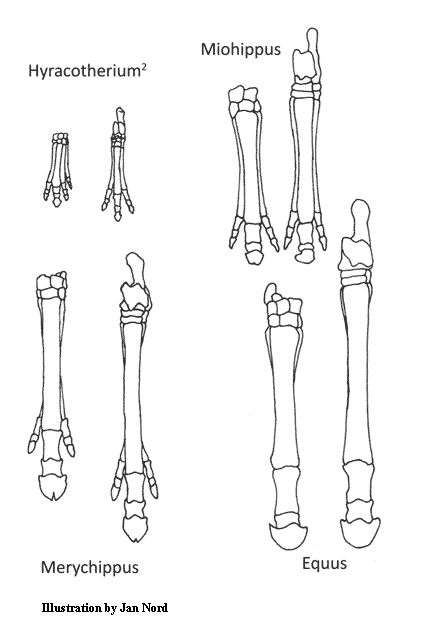 Obrázek 2. Nohy koní pokládané za důkaz evoluce. Levá noha v každém zobrazeném páru je nohou přední a pravá noha nohou zadní6.
Obrázek 2. Nohy koní pokládané za důkaz evoluce. Levá noha v každém zobrazeném páru je nohou přední a pravá noha nohou zadní6.
Po roce 1989 byl zpochybněn monofyletický charakter Hyracotheria9
Roku 1992 byl rod Hyracotherium překlasifikován na pět zvířat patřících do různých čeledí, ze kterých jen u jedné skupiny byla uznána jakási příbuznost s koňmi15. Ještě modernější bádání překlasifikovalo tato zvířata do deseti různých rodů a nejméně tří čeledí, přičemž o mnohých z nich se soudí, že neměla nic společného s řadou koně, nýbrž že byla podobná např. tapírům (čeleď Tapiromorpha)9. Jeden druh Hyracotheria (angustidens) byl přejmenován na Eohippus a všem dalším druhům Hyracotherií až na jeden byla dána nová rodová jména. Ono jediné zvíře vystupující nadále pod jménem Hyracotherium (leporinum) není už v řadě koně, nýbrž je považováno za blízkého příbuzného Palaeotherií, která připomínají tapíry a nosorožce.
„Rané“ koně byli zachováni ve vrstvách ze stejného evolučního období jako někteří „pozdější“ koně
Hyracotherium/Eohippus a Orohippus se například skutečně objevují ve fosilním záznamu v téže době jako Epihippus. Mesohippus a Miohippus se objevují společně s Merychippem a Parahippem. Téměř všichni další koně (možná s výjimkou jednoho či dvou) – Parahippus, Merychippus, Pliohippus, Equus a možná také Miohippus – jsou zastoupeni v téže době, ve stejném časovém úseku, ve kterém jsou nalézáni jako fosilie16. (Ale zejména v novějších evolučních schématech byla dána odlišná jména velmi podobným zvířatům, což činí dojem evoluce, a zároveň přináší slávu svým objevitelům; viz příklady v Froehlich 20029 a MacFadden 20054). Fosilie Hyracotheria (sic) byly nalezeny též ve velmi vysokých vrstvách (pliocén), ale tyto nálezy byly odmítnuty jako přemodelované (tj. erodované a uložené v pozdějších vrstvách) navzdory faktu, že geologická pozorování neprokázala žádné známky porušení17. Takže skutečnost, že většina koní žila skoro ve stejné době, podkopává teorii o jejich údajné evoluci.
„Přechodné“ formy mezi koňmi se zuby určenými pro spásání listí (Parahippus) a koňmi se zuby pro spásání trávy (Merychippus) jsou vzácné13
 Obrázek 3. Stavba zubů u koní živících se listím (dva druhy vlevo) a trávou (dva druhy vpravo)7.
Obrázek 3. Stavba zubů u koní živících se listím (dva druhy vlevo) a trávou (dva druhy vpravo)7.
Zuby koní živících se listím mají uzavřené, velmi úzké kořeny s malými otvory pro nervy a zásobování krví; tzn., že jde o zuby, které se opotřebovávají s tím, jak zvíře stárne. Zuby koní živících se trávou mají otevřený kořen s mnoha krevními cévami, které zásobují zuby spoustou živin, takže mohou dorůstat po celý život zvířete; říká se tomu hypsodoncie, což znamená zuby s vysokými korunkami. Tato změna struktury zubů z bunodontních (nízké korunky se zaoblenými hroty) v hypsodontní (vysoké korunky) není pouhou údajnou „mikroevolucí“, nýbrž dokonalou změnou designu, i když se to lidem neobeznámeným s konstrukcí zubů nemusí snad jevit jako něco tak převratného18. Neexistují důkazy jakékoli změny od jedné struktury zubů ke druhé, i když se o ní někteří autoři zmiňují19. Některá zvířata se živila jak trávou, tak listím3, 4, což nám však nepomůže vysvětlit transformaci jednoho druhu zubů v jiný.
Tři zcela rozdílná zvířata
Zvířata, která byla vykládána jako různé druhy koní, můžeme proto, budeme-li mít na paměti shora uvedená fakta, snadno určit jako tvory patřící ke třem zcela rozdílným stvořeným druhům zvířat, a nikoli jako různé přechodné formy ve vývoji koně, které se údajně vyvinuly z jednoho a téhož původního předka. Ony stvořené druhy, s vyloučením všech zástupců Hyracotheria, kteří byli přesunuti do nových čeledí, by proto měly více či méně odpovídat následujícím třem skupinám (pamatujme, že v dalším výčtu nejsou uvedeni všichni nově pojmenovaní koně ani všichni zástupci vedlejších skupin):
- Eohippus (a mnoho fosilií, které byly dříve označovány jako Hyracotherium, ale jsou zařazeny do čeledi Equidae s novými rodovými jmény9), Orohippus a Epihippus.
- Mesohippus, Miohippus, jistí zástupci rodu Parahippus, a patrně většina koní řazených do rodokmenů těchto tří skupin. (Řada koně byla přestavěna a bylo doplněno mnoho nových rodů; např.větve Neohipparion, Nannipus a Hipparion byly přesunuty hned vedle Parahippa a odsunuty od Merychippa4, na rozdíl od obrázku 1, takže si nemůžeme být jisti, zda je klasifikace/rozdělení do skupin u všech fosilií správné. Ale koně větvící se z Merychippa na obrázku 1 jsou stále řazeni do podčeledi Equinae, a jsou proto zde uspořádáni do skupiny následující, třetí. Všemi těmito podrobnostmi se však v tomto článku nemůžeme zabývat).
- Merychippus a koně větvící se z této skupiny včetně Pliohippa a všech pozdějších koní (včetně větví rodu Hipparion). (Všimněte si, že v nových revizích evoluce koně existují dva různé rody se jménem Merychippus: I a II. Merychippus je proto považován za polyfyletického, tzn., že se soudí, že se vyvinul dvakrát. Tyto dva rody byly umístěny do odlišných evolučních linií. Rod I je na původním místě vedoucím k Equus, jak vyplývá z většiny diagramů evoluce koně. Rod II byl odstraněn z linie vedoucí k Equus – je považován za současníka Parahippa po většinu času, kdy žil – a je považován za předka vývojových větví začínajících u Hippariona, jak to popisuje MacFadden 2005)4.
 Obrázek 4. Dva „koně“, Neohipparion (vpravo) a Miohippus (vlevo) z Přírodovědného muzea v Los Angeles.
Obrázek 4. Dva „koně“, Neohipparion (vpravo) a Miohippus (vlevo) z Přírodovědného muzea v Los Angeles.
Zvířata ve skupině 3 jsou všechna řazena do téže podčeledi – Equinae20. I když také Cavanaugh et al.10 objevili, že by se fosilní zvířata mohla řadit do podčeledí, zavrhli nakonec tohle zjištění a místo toho zkonstruovali vlastní rodokmen koňské evoluce. Nebylo by těžké vytvořit rodokmen podobný, kdybychom prostě seřadili jakékoli množství nepříbuzných žijících zvířat do řady od malého k velkému (obrázek 5).
Žádná evoluce koně se nekonala
Hypotéza Cavanaugha, Wooda a Wiseho14, že řada koně (včetně rodu Hyracotherium) svědčí o skutečné (popotopní) „mikroevoluci“ (či lineární/progresivní variaci), je s ohledem na shora uvedené skutečnosti neudržitelná, jelikož v evoluci koně neexistuje žádný pokrok (kromě snad lokálních rozdílů) a z dat vyplývá směs různých, koni podobných zvířat. Navíc se článek Cavanaugha et al.14 opíral hlavně o statistická data z jednoho zdroje z roku 1989 (a některé diskuze z novějších kreacionistických časopisů) a nevyhodnotil jednotlivé nálezy Hyracotheria. Rovněž Froehlichův článek9, který zavedl novou nomenklaturu pro všechny druhy Hyracotherií, byl zveřejněn v únoru 2002, zhruba rok před uzávěrkou článku Cavanaugha et al. 2003 v rámci ICC14. Tyto nejasnosti ohledně nálezů Hyracotheria nebyly zohledněny ani v článku Wooda z roku 200821, ačkoli se Wood odvolává na knihu MacFaddena z roku 199222, který konstatoval, že Hyracotherium nebylo jediné zvíře, nýbrž několik rodů patřících do různých čeledí. Whitmore a Wise dokonce roku 2008 používají Hyracotherium ke stanovení doby jeho výskytu krátce po Potopě, a toto zvíře, které nebylo koněm, je uvedeno jako první zástupce v řadě koně23.
 Obrázek 5. Zleva doprava antilopa losí (oryx), pakůň běloocasý (gnu), lesoň pestrý (bušbok), gazela a antilopa dikdik. Dokonce i dnes žijící zvířata lze seřadit do hypotetické evoluční řady, jelikož se proměny kostry uvnitř jedné skupiny zvířat často překrývají s proměnami v jiných skupinách v rámci téže čeledi. To však nedokazuje, že se jakékoli konkrétní zvíře vyvinulo v jiné.
Obrázek 5. Zleva doprava antilopa losí (oryx), pakůň běloocasý (gnu), lesoň pestrý (bušbok), gazela a antilopa dikdik. Dokonce i dnes žijící zvířata lze seřadit do hypotetické evoluční řady, jelikož se proměny kostry uvnitř jedné skupiny zvířat často překrývají s proměnami v jiných skupinách v rámci téže čeledi. To však nedokazuje, že se jakékoli konkrétní zvíře vyvinulo v jiné.
Froehlich9, který zcela přejmenoval většinu druhů Hyracotheria a zařadil je do různých rodů a čeledí, použil statistiku, ale vyjádřil i svůj kritický postoj ke způsobu, jakým v tomto případě může být statistika zneužita. V každém případě však nelze užít statistiku k hodnocení designu či tam, kde je k dispozici jen omezený objem dat (což jsou v těchto případech většinou zuby a čelisti), aby se zjistilo, jak údajně probíhala evoluce, jak to učinili svrchu citovaní autoři9, 14. Statistická analýza nebere v tomto případě v úvahu funkci či hotové/naplánované živé entity, nýbrž může jen porovnávat malé rozdíly (viz též zásadnější kritiku u Froehlicha)9. V tomto případě se většina statistické analýzy opírala o malé rozdíly v zubní sklovině/struktuře zubů a v čelistech a velmi málo byly zohledněny ostatní části těla. To samozřejmě činí interpretaci dat neobjektivní podobným způsobem, jako kdybychom například provedli statistické zhodnocení na základě 75 rozdílů ve vnějším vzhledu očí chobotnic a lidí – z takové analýzy by patrně vyplynulo, že jsme se vyvinuli z chobotnic.
Ačkoli je snadné rozebírat a kritizovat jednotlivé nálezy či jediné místo, kde byly fosilie nalezeny, v souladu se všemi daty, která jsou k dispozici, se zdá, že jsou zde tři skupiny zvířat, které bezprostředně odpovídají skupinám – podčeledím koňovitých, a vypadá to, že pouze v podčeledi Equinae jsou zastoupeni koně. Diskuze o popotopní situaci i situaci v době Potopy vycházející z evoluce koně a probíhající např. u Cavanaugha et al. 200314 a Wooda 200821 se proto musí opírat o jiná kritéria než je údajná popotopní „mikroevoluce“ koně jako důsledek měnícího se prostředí, jak o tom obvykle hovoří evolucionisté (viz odlišná kritéria pro okolnosti kolem Potopy u Oarda 2007)24. Neexistovala rovněž žádná konkrétní území, kde by tato zvířata mohla žít, jen rozsáhlé pouště – většinu fosilií nalézáme v sedimentárních nánosech svědčících o tom, že je nashromáždila Potopa, ale neexistují důkazy o rostlinném pokryvu, který by mohl poskytovat potravu velkým stádům zvířat, ani o půdě v pravém smyslu slova25. Nemáme rovněž důkazy o změnách prostředí, na které poukazují evolucionisté i Cavanaugh et al.14 a Wood21 vycházející ze spekulativních výkladů.
V případu koně by to mohly být tělesné rozměry, které určily, jak rychle zvířata utonula, byla unesena proudy a pohřbena, a pak občas obnažena erozí a znovu uložena, během Potopy či během procesů následujících bezprostředně po Potopě. Zřejmě k tomu došlo předtím, než se stala území kontinentů opět obyvatelnými a znovu se na nich zabydlila živá zvířata. Malá zvířata s podobnou konstrukcí se obvykle rozkládají a klesnou ke dnu rychleji než velká zvířata a menší kosti jsou také snadněji unášeny proudy poté, co dosáhly dna. Rovněž mohla být během popotopních katastrof žijící zvířata pohřbena společně s přemodelovanými, mrtvými, nefosilizovanými či částečně fosilizovanými zvířecími ostatky, pohřbenými během Potopy.
Závěr
Studium fosilních koní přináší poznatky o přinejmenším třech skupinách zvířat v rámci čeledi koní Equidae, a navíc o některých nepříbuzných zvířatech jako třeba tapírech. Ony tři skupiny koním podobných zvířat blízce korespondují s různými podčeleděmi čeledi Equidae a bylo by možno je považovat za tři zvláštní stvořené druhy. Většina z těchto odlišných druhů žila (či vlastně byla pohřbena!) zhruba ve stejné době a nevykazuje žádné velké progresivní změny, které by svědčily o evoluci koně, pouze všeobecný zvyšující se tělesný vzrůst.
Nikdo nevysvětlil, jak se asi mohly vyvíjet nové, specializované druhy zubů, a zdá se, že jde spíše o případ inteligentního plánu a nikoli o „mikroevoluci“ (variace v rámci stvořeného druhu, jak o tom píší různí kreacionisté); ani se nejedná o „makroevoluci“ (nových druhů organizmů, jak tvrdí evolucionisté).
Hypotéza Cavanaugha et al. (2003)14 o intrabaraminické proměnlivosti všech zvířat patřících do čeledi Equidae (či zvířat, která řadí do Equidae, třebaže i evolucionisté řadí některá z nich do jiných čeledí) není patřičně podpořena důkazy, které jsou k dispozici, a měla by proto být opuštěna.
Dodatek
Podle Juliana Huxleyho (možná jednoho z nejprominentnějších evolucionistů minulého století) bylo potřeba přinejmenším jednoho milionu pozitivních mutací, aby se vyvinul moderní kůň. Huxley věřil, že existuje maximální počet jedné pozitivní mutace na každých 1 000 mutací ostatních. Pomocí těchto hodnot Huxley propočítal, že pravděpodobnost, že se kůň vyvine z jediného jednobuněčného organizmu, je 1 ku 103 000 000. Věřil však, že přírodní výběr by byl schopen tento problém vyřešit26. Tato víra mu však nakonec nepomohla, ani nepomůže kterémukoli jinému evolucionistovi, jelikož je tato kalkulace založena na vzniku pozitivních mutací ještě předtím, než by snad začal fungovat přírodní výběr. Kdyby se totiž všechny elektrony ve vesmíru (zhruba 1080) účastnily 1012 reakcí každou sekundu, nenastalo by ani během 30 miliard let, která evolucionisté stanovili jako horní hranici stáří vesmíru, více než cca 10110 možných interakcí – což je stále daleko od Huxleyho propočtu1.
Odkazy
- This article is partly based on an English translation (by Rudi Arbella) of Molén, M., Vårt ursprung? (Our Origin), XP-media, Haninge, 2000.
- The name Hyracotherium is no longer in use for any member of the family Equidae, but the fossils of this genus which are still classified in Equidae have been given seven new genus names: Sifrhippus, Minippus, Arenahippus, Xenicohippus, Eohippus, Plilolophus and Protorohippus.
- Some horses with teeth designed to eat grass probably mostly ate leaves and other soft plant parts. See: Morell, V., Dietary data straight from the horses mouth, Science 283:773, 1999; and MacFadden, B.J. et al., Ancient Diets, Ecology, and Extinction of 5-Million-Year-Old Horses from Florida, Science 283:824–827, 1999.
- MacFadden, B.J., Fossil horses—evidence for evolution, Science 307:824–827, 2005.
- Simpson, G.G., Horses, Oxford University Press, New York 1951.
- The picture is based on drawings from: Steinmann, G. and Döderlein, L., Elemente der Paläontologie (Elements of Paleontology), Wilhelm Eng-elmann Publishing, Leipzig, 1890; Osborn, ref. 11; and Scott, ref. 12.
- The picture is based on drawings from: Steinmann and Döderlein, ref. 6; Osborn, ref. 11; Mutvei, H. and Stuenes, S., Paleontologi: Kompendium för grundkurs i paleontologi (Paleontology: Compendium for Basics in Paleontology), Uppsala University, Institute of Paleontology, Uppsala, 1983; and Granger, W., A revision of American Eocene horses, Bulletin of the American Museum of Natural History 24:221–264, 1908.
- MacFadden, B.J., Cladistic analysis of primitive equids, with notes on other perissodactyls, Systematic Zoology 25:1–14, March 1976; and Simpson, G.G., Horses, Oxford University Press, New York, 122–123, 203, 1951.
- Froehlich, D.J., Quo vadis eohippus? The systematics and taxonomy of the early Eocene equids (Perissodactyla), Zoological Journal of the Linnean Society 134:141–256, February 2002.
- Abel, O., Paläobiologie und Stammesgeschichte (Paleobiology and the History of Phylogeny), Gustav Fisher Publishing, Jena, pp. 284–290, 1929; and MacFadden, B.J. et al., Sr-isotopic, paleomagnetic, and biostratigraphic calibration of horse evolution: evidence from the Miocene of Florida, Geology 19:242–245, 1991.
- Osborn, H.F., Equidae of the Oligocene, Miocene, and Pliocene of North America: iconographic type revision, Memoirs of the American Museum of Natural History 2:74–75, 1918. One example is that one of the most “primitive horses”—Para-hippus cognatus—supposedly evolved as one of the last species, while one of the least primitive— coloradensis—evolved at the same time as the first species of Parahippus.
- Scott, W.B., A History of Land Mammals in the Western Hemisphere, 2nd, Macmillan Publ. Co., NY, p. 409, 1937.
- In the Parahippus group there are some findings that are assumed by some researchers to be intermediate forms between Parahippus and Merychippus (e.g. with reference to pictures in Osborn, ref. 11, where some of the least “evolved” “horses” originated amongst the last ‘horses’; see pp. 74–75). These findings only consist of teeth and parts of jaws that can be difficult to tie to a certain skeleton. Hence, these teeth and jaws could therefore be sorted into Parahippus and Merychippus, rather than any intermediate form between these two animals.
- Cavanaugh, D.P., Wood, T. and Wise, K.P., Fossil equidae: a monobaraminic, stratomorphic series; in: Walsh, R.E. (Ed.), Proceedings of the Fifth International Conference on Creationism, Creation Science Fellowship, Pittsburgh, PA, p. 143–149, 2003.
- MacFadden, B.J., Fossil Horses, Cambridge University Press, Cambridge, pp. 92–103, 1992. See also Scheven, J., Mega-Sukzessionen und Klimax im Tertiär: Katastrophen zwischen Sintflut und Eiszeit (Megasuccessions and Climax in the Tertiary: Catastrophes Between the Flood and the Ice Age.), Wort und Wissen, Neuhausen, 1988.
- Scott, ref. 12, p. 408 and Osborn, ref. 11, p. 74; Barnhart, W.R. A Critical Evaluation of the Phylogeny of the Horse, ICR, 1987; MacFadden, ref. 15, pp. 255–257; Storer, J.E. and Bryant, H.N., Biostratigraphy of the Cypress Hills Formation (Eocene to Miocene), Saskatchewan, Journal of Paleontology 67:660–669, 1993; Gould, S.J. and Eldredge, N., Punctuated equilibrium comes of age, Nature 366:223–227, 1993; and Scheven, ref. 15.
- Barnhart, ref. 16, pp. 140–143
- Personal communication with Dr Jack Cuozzo. Return to text.
- Sarfati, J., The non-evolution of the horse, Creation 21(3):28–31, 1999.
- Gromova, V.I., Mammals in: Fundamentals of Paleontology: A Manual for Paleontologists and Geologists of the USSR, vol. 13, Orlov, Y.A. (Ed.), Jerusalem, English translation, Israel Program for Scientific Translations, p. 396, 1968.
- Wood, T., Horse fossils and the nature of science, 2008; answersingenesis.org/articles/am/v3/n4/horse-fossils.
- MacFadden, ref. 15, pp. 92–103.
- Whitmore, J.H. and Wise, K.P., Rapid and early post-Flood mammalian diversification evidenced in the Green River Formation; in: Walsh, R.E. (Ed.), Proceedings of the Sixth International Conference on Creationism, Creation Science Fellowship, Pittsburgh, PA, pp. 449–457, 2008.
- Oard, M.J., Defining the Flood/post-Flood boundary in sedimentary rocks, Journal of Creation 21(1):98–110, 2007.
- Molén, M., Mountain building and continental drift; in: Walsh, R.E. (Ed.), Proceedings of the Third International Conference on Creationism, Creation Science Fellowship, Pittsburgh, PA, pp. 353–367, 1994.
- Huxley, J., Evolution in Action, Chatto and Windus, London, pp. 47–48, 1953.