
Hlavním problémem neodarwinismu je chybění fosilních důkazů pro evoluci členovců
Jerry Bergman
Creation Research Society Quarterly, roč. 43, č. 4, březen 2007, str. 222 – 228:
Jerry Bergman, Ph.D., Department of Biology Northwest State College, Archbold, OH 43502.
Přeložil M. T.
Souhrn
Zachované fosilie kmene členovci jsou hlavním problémem pro neodarwinistický evolucionismus. Nikdy nebyl objeven žádný jasný fosilní důkaz pro evoluci členovců, ačkoli byly nalezeny milióny fosilií členovců. Nálezy jako celek jasně ukazují statický stav čili neměnnost druhů, nikoli progresívní evoluci od jednoho řádu k jinému. Velká část všech žijících i vymřelých druhů zvířat patří do kmene členovců, a naprostý nedostatek fosilních důkazů pro makroevoluci celého tohoto kmene svědčí ve prospěch teorie o Stvoření.
Úvod
Členovci jsou nejrozsáhlejším a nejdiferencovanějším kmenem v celé živočišné říši a zahrnují přes jeden milión různých druhů (Giribet a Ribera, 2000). Asi 78% všech popsaných žijících druhů zvířat jsou členovci (Wills, 2001). Nalézáme je v tekoucích sladkých vodách, v moři i na suchozemských stanovištích po celém světě (Wills et al., 1997). Členovci nejsou pouze největším kmenem v živočišné říši, ale jsou též jediným kmenem bezobratlých, jenž zahrnuje vodní, suchozemské i vzdušné zástupce. Zahrnuje trilobity, korýše, pavoukovce, stonožky, hrotnatce (ostrorepy, nezahrnuté do tohoto článku – viz Helder, 1997), a šestinožce (hmyz a tři malé řády hmyzovitých zvířátek, rovněž nezahrnutých do tohoto článku – viz Bergman 2004 a Manning 2003). Jsou tak rozdílní, že někteří entomologové se ptají, zda mohou pocházet ze společného předka či nikoli. Běžně se učí, že ze společného předka pocházejí (Manton, 1973). Lammerts (1974) vyvodil ze svého studia entomologie závěr, že
„pro mne bylo zdrojem stálého úžasu, jak mohl být tak pozoruhodný soubor tvorů (jež nalézáme v řádu brouci) kdy považován za potomky jediného společného předka. Jeden z mých profesorů, Dr. E. C. VanDyke, světová autorita na tuto skupinu, byl často stejně jako já tímto předpokladem ohromován (str. 125).
Členovci jsou definováni jako „bilaterálně symetrická článkovaná zvířata s charakteristickým tuhým chitinizovaným obranným exoskeletonem ohebným pouze v kloubech; k růstu dochází ekdyzí“ (Tootill, 1988). Růst ekdyzí (často zvaný línání) vyžaduje pravidelné svlékání kutikulového exoskeletonu a růst exoskeletonu nového a většího. Každý článek těla členovců obsahuje většinou pár dlouhých přívěsků modifikovaných tak, aby sloužily různým účelům. Célom členovce je dlouhá dutina naplněná tekutinou umístěná mezi stěnou břišní a stěnou tělní.
Célom členovců kromě hmyzu funguje jako hydrostatická kostra, a jeho velikost je ve srovnání s célomem hmyzu menší. Hlavní tělní dutinou členovců je célom naplněný hemolymfou (krvomízou), struktura známá jako hemocoel. Členovci mají též břišní nervovou pásku, pár mozkových ganglií a pár uzlin v každém článku (Tootill, 1988). Jsou také „jedinými bezobratlými, kteří vykazují pevnou, individualizovanou formu tukové tkáně“ (Kaufmann, 1977, str. 214).
Zachované fosilie členovců obecně
Darwin spatřoval ve fosilních nálezech hlavní problém své teorie a očekával, že čas zaplní mezery nyní tak slavných „chybějících spojovacích článků“. Většina fosilních důkazů užívaných v pokusu podpořit neodarwinismus sestává z fragmentů kostí obratlovců, jež vyžadují značné interpretační úsilí, což vedlo k velkým sporům mezi paleontology specializovanými na obratlovce. Z odhadovaných 1,7 miliónu známých druhů zvířat je však plných 95% bezobratlých, a většinu z nich tvoří právě členovci (Barnes, 1987). Existuje velké množství fosilií mnoha bezobratlých včetně členovců – prostě milióny. Mnoho z těchto fosilií členovců vykazuje skvěle zachovanou morfologii těla (Eldredge, 2000). A dále, „některé z nejstarších, nejpozoruhodnějších a nejinformativnějších mnohobuněčných fosilních živočichů jsou …členovci“ (Wills, 2001, str. 187). Nalézáme je v hojném počtu v horninách označovaných jako paleozoikum (prvohory), mezozoikum (druhohory) i kenozoikum (třetihory a čtvrtohory). Je jich tam víc než fosilií kterékoli jiné skupiny organismů. Jsou zachovány i případy interakcí rostliny a členovce z tak vzdálené doby jakou je zmiňované paleozoikum (Chaloner et al., 1991).
I když jsou s rozsáhlými časovými úseky připisovanými těmto předpokládaným geologickým věkům spojeny značné problémy a jsou často zpochybňovány, budeme se jimi v naší studii řídit jako odkazovým rámcem. Larvy členovců dlouhé necelý milimetr byly nalezeny v horninách svrchního kambria (Anderson, 1998, str. 434). Bohaté nálezy fosilií členovců byly učiněny v takových vynikajících ochranných látkách jako jantar, asfalt či led. Tam jsou často zachovány výborné detaily i těch nejnepatrnějších tělesných struktur jako jsou oči a chloupky (Gayrard-Valy, 1994; Alonso et al., 2000). Jiným důvodem pro bohaté nálezy těchto fosilií je fakt, že někteří členovci včetně trilobitů, rakovců a lasturnatek, mají kostry vyztuženy vápenatými solemi (Wills, 2001).
Suchozemské členovce nalézáme nejdříve před spodním devonem v evolučním časovém schématu (Edgecombe, 1998a, str. 174). Tyto výborně zachované fosilní nálezy umožňují přesná srovnání starých a moderních forem členovců, jež lze použít při posuzování věrohodnosti darwinismu. Pro příklad viz Delclose et al., 2000. Budd (1997) uzavřel svá pozorování slovy, že „však není zdaleka jasné, co tyto fosilie znamenají jak pro klasifikaci členovců tak pro ranou evoluci kmene: ani jeden názor nezískal univerzální podporu“, protože zachované fosilie nedokládají jasně makroevoluci (str. 125).
Dráhy zanechané plazícími se zvířaty a vyvrtané otvory po členovcích jsou též běžné a důležité ve fosilních nálezech, zejména nemarinních členovců (Donovan, 1994, str. 200). Dále byly nalezeny též sítě starých či fosilních pavouků jakož i vosí, včelí a mravenčí hnízda.
Velkým problémem při stanovování evolučních vztahů kmene členovci je nedostatečná shoda na tom, co přesně rozumíme pod definicí kmene či jiné taxonomické kategorie (Meglitsch a Schram, 1991). A ještě větším problémem je určit, zda oni tvorové, jež dnes považujeme za členovce, patří skutečně do jediného kmene členovci (Anderson, 1998). Spory se vedou i v tak zásadní otázce jako je ta, zda fosilní nálezy stačí k tomu, abychom stanovili fylogenetický rodokmen. Například:
Fosilní nálezy korýšů, často v učebnicích označované jako chudé, jsou ve skutečnosti dobré. Všechny čtyři třídy korýšů zanechaly fosilie… Nálezy jsou ve skutečnosti tak četné, že omezený prostor naší studie nám nedovoluje odpovídající diskusi. (Meglitsch a Schram, 1991, str. 490).
Ačkoli je známa „hojnost“ fascinujících fosilních členovců (Meglitsch a Schram, 1991, str. 368), nálezy poskytují málo důkazů jak pro jejich původ tak pro jejich vývoj v makroevoluci. Výsledkem je, že existuje „handrkování nad nesčetnými hypotézami a scénáři“ pro
„evoluci členovců. Zdá se, že každá autorita na tomto poli má vlastní verzi. Neexistuje ani shoda na tom, zda máme co dělat s jediným kmenem členovci“ (Meglitsch a Schram, 1991, str. 369).
Podstatným problémem pro darwinismus je fakt, že některé z nejstarších forem živočišného života nalézané coby fosilie jsou též jedněmi z nejsložitějších (Eldredge, 2000, str. 42). O faktech týkajících se fosilií členovců se mnoho nediskutuje, zato se vedou urputné spory o důsledcích těchto faktů (Morris, 2000: Hallam, 1997).
Aguinaldo et al., (1997) konstatoval, že
členovci představují nejrozmanitější skupinu zvířat, avšak přestože nálezy jejich fosilií jsou bohaté a máme za sebou jedno století studia, jejich fylogenetické vztahy zůstávají nejasné. Taxony dříve považované za sesterské skupiny členovců zahrnují kroužkovce, drápkovce, želvušky a jiné, ale hypotézy o jejich možných fylogenetických vztazích si odporují. (str. 489)
Bylo nalezeno tolik fosilií, že si někteří paleontologové myslí, že „rané pozemské ekosystémy ovládali malí členovci“ (Shear et al., 1996, str. 555). Navzdory faktu, že „fosilie jsou zachovány v perfektním stavu, existuje v nálezech členovců mnoho nápadných mezer“ a, jako důsledek, „vztahy hlavních skupin členovců obklopuje nesmírná kontroverze“ (Wills, 2001, str. 188, 190). Některé uvažované fylogenetické vztahy mezi členovci můžeme najít v pracích Valentineho (1989) a Edgecomba (1998a).
Trilobiti

Trilobity oplývají v hojné míře zejména vrstvy označované jako kambrium a silur. Dosud bylo popsáno asi 65 různých rodů a přes 100 druhů trilobitů (Margulis a Sagan 2002, viz str. 180). Trilobiti mají ploché, oválné tělo rozdělené podélně do tří laloků – odtud jejich název. Jako všichni ostatní členovci mají i trilobiti hlavu, hruď a zadeček. Podobně jako rasy psů jsou známy nesčetné variety trilobitů, všichni však mají tři laloky a chitinizovaný exoskeleton (Sherwin a Armitage, 2003). Trilobity snadno poznáme, protože mnoho jejich rysů (jako třeba celkové uspořádání organizace končetin) je pro ně ve srovnání s jinými skupinami členovců specifické (Eldredge, 1977, viz str. 327).
Trilobiti jsou zachováni mimořádně dobře, zčásti proto, že jejich tvrdý karapax z uhličitanu vápenatého (pokrývka těla) je vysoce odolný vůči rozkladu (Margulis a Sagan, 2002). Z tohoto důvodu jsou jejich fosilní nálezy jedny z nejlepších pokud jde o rostliny i živočichy. Trilobity nalézáme v „trilobitových ložiscích“ na mnoha místech Země (Meister, 1968). Z toho důvodu jsou také zdaleka jednou z nejpozorněji studovaných ze všech skupin bezobratlých (Clarkson, 1986). Trilobiti jsou „většinou zachováni coby chitinizované desky“ či jako skořápky a v některých případech je „jejich současná kokila tak dobře zachována, že stále můžeme pozorovat podstatné znaky na povrchu jejich těl“ (Twenhofel a Shrock, 1935, str. 433).
Zejména oči trilobitů byly rozsáhle studovány stejně jako jejich četné párované přívěsky. Naštěstí jsou tato zvířata zachována ve fosilních nálezech velmi detailně (viz Levi-Setti, 1993, viz str. 45 – 54 pro fotografie SEM, jež ukazují tyto podrobnosti). Oči trilobitů tvoří obrovské, složité, složené facetované struktury, jež někdy tvoří souvislý pruh přes celý lem jejich čela (Prokop, 1995). Trilobití oči se podobají očím moderního hmyzu a byly patrně nejméně stejně tak složité. Vzhledové prvky jako jsou uvedené struktury silně podporují myšlenku, že jde o dílo inteligentního Tvůrce (Sherwin a Armitage, 2003; DeYoung, 2002).
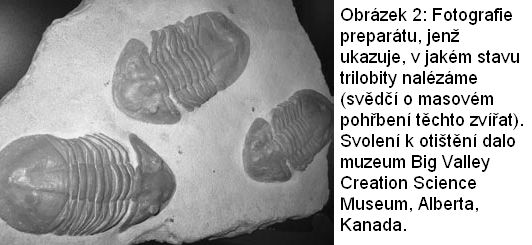
Trilobiti jsou důležití jak pro neodarwinistickou tak pro kreacionistickou pozici (Cook, 1968). Eldredge (2000) konstatuje, že trilobiti „jsou zrovna tak důležité příklady evoluce jako všechny ostatní, které znám“, vykazují však pouze mikroevoluci (str. 122). Byly jich nalezeny již milióny ve výborném stavu, který dovoluje přesnou rekonstrukci jejich historie, ale nedokládá nikterak makroevoluci. Namísto toho dokládají fosilní nálezy „relativně náhlé, překotné vynoření se trilobitů i jiných složitých forem živočišného života počátkem kambria“ (Eldredge, 1977, str. 44).
Jak poznamenal Clarkson (1986), „o předcích trilobitů nevíme téměř nic. Společně s ostatními členovci můžeme předpokládat, že trilobiti snad vzešli ze stejných předků jako kroužkovci…ale jde o pouhou spekulaci“ (str. 330). Mnoho badatelů soudí, že trilobiti jsou nejblíže příbuzní s korýši. Většina druhů trilobitů existovala zároveň ve stejné době, což vylučuje předpoklady nějakých jasných evolučních trendů v rámci existujících fosilií.
Rozsáhlé studium jejich fosilií ukázalo, že trilobiti „ve svých druzích a poddruzích mají tendenci zůstávat relativně nezměnění…během rozpětí svého výskytu v ložiscích“ a že existuje „méně dlouhodobých ´evolučních trendů´ v rozsahu linií ´prokázaných´ u trilobitů než je tomu u mnoha jiných skupin“ (Eldredge, 1977, str. 309, 316). Ve fosilních nálezech se však přece jen setkáváme s určitým zakrňováním (devolucí), jako je ztráta očí (Whittington, 1992).

Fosilní nálezy neposkytují důkazy relativně rychlé evoluce následované stagnací a pak další evolucí. Místo toho vykazují „náhlý výskyt různých druhů trilobitů ve spodním kambriu a nahrazení těchto raných skupin novými během přechodu z kambria do ordoviku“ (Whittington, 1992, str. 84). A dále, nebyly nalezeny „žádné důkazy (jako třeba přechodná série fosilií)“ k překlenutí propastí oddělujících od sebe různé druhy trilobitů (Whittington, 1992, str. 85, a též tabulka str. 86). Chybění přechodných forem je v souladu s modelem původu života Stvořením.
Spekulace o evoluci trilobitů činěné ve velkém paleontology je normou, což vedlo Whittingtona ke konstatování, že „neexistuje nedostatek ani interpretací ani spekulací“ (1992, str. 85). Eldredge (2000) se pokusil zdůvodnit nedostatek fosilních důkazů pro evoluci trilobitů uchýlením se k modelu přerušované rovnováhy – teorii, že evoluce probíhala po relativně krátké časové úseky následované dlouhými periodami stagnace (viz str. 44). Eldredge a Gould vlastně vyvinuli svou teorii přerušované rovnováhy speciálně proto, aby vysvětlili nedostatek fosilních důkazů pro evoluci trilobitů (Eldredge, 1977, 2000; Gould, 1977)
Velká množství trilobitů z neznámých příčin vymřela na konci kambria, a z fosilních nálezů se dovídáme, že příval nových druhů se objevil v horninových formacích připisovaných dalšímu období, ordoviku. Avšak „po tomto velkém boomu trilobitích kmenů nového ustrojení ve spodním ordoviku se vynořilo jen velmi málo zcela nových organizačních vzorců…trilobiti jako takoví zůstali konstruováni na témže archetypálním plánu definovaném v nejspodnějším kambriu“ (Clarkson, 1986, str. 331). Všichni trilobiti vymřeli, jako ostatně mnoho dalších zvířat, v období nazývaném evolucionisty velké permské vymírání.
Korýši
Podkmen korýšů zahrnuje přes 35 000 druhů po celém světě, zejména v tekoucích vodách a v moři. Pro příklady korýšů viz obrázek 3. Korýši mají ze všech členovců nejrozmanitější odchylky ve stavbě těla mezi sebou a zahrnují garnáty, kraby, humry, vilejše, svinky, perloočky a buchanky (jako třeba dafnie=hrotnatky) a kapřivce (Giribet a Ribera, 2000, str. 220). Mají pět párů pohybových přívěsků, z nichž první dva mohou tvořit klepeta sloužící obraně a manipulaci s potravou. Mají též komplexní, dobře vyvinuté oči (Hamilton, 1986). Nejstarší fosilie korýšů se datují až do časů spodního kambria. Anderson poznamenává, že ačkoli
jsou fosilní nálezy docela bohaté u mnoha skupin korýšů, nepoučují nás o jejich původu, a existuje zásadní debata o tom, jak vypadala primitivní či předkovská těla prvních korýšů. Současný názor dává přednost dlouhému tělu s mnoha podobnými segmenty trupu, dvěma páry biramózních (přívěsky členovců, jež mají dvě ramena) antén a naupliovou (larvální stadium charakterizované třemi přívěsky) larvou, ale vědci jsou rozděleni v otázce, zda trup měl biramózní či polyramózní (přívěsky členovců, jež mají více než dvě ramena) plovací přívěsky (1998, str. 316, závorky přidány).
Jiní badatelé nesouhlasí a argumentují tím, že se korýši vyvinuli z jakéhosi spirálního červa či dávného ploštěnce (Willmer, 1990, viz str. 298 – 299). Fylogenetické vztahy korýšů jsou komplikované a existuje málo shody mezi vědci (Giribet a Ribera, 2000, viz str. 220).
Pavoukovci (klepítkatci)
Pavoukovci (pavouci, štíři, solifugy, roztoči a klíšťata) mají čtyři páry přívěsků – nohou. Pavouci jsou skupinou pavoukovců, která je sedmá co do počtu druhů mezi všemi zatím popsanými a odhadnutými zvířaty. Po celém světě běhá asi 30 000 až 40 000 druhů pavouků, a to od Arktidy až po pouště (Williams a Goette, 1997, viz str. 3 – 4). Jedním z nejobávanějších pavouků je tarantule, velký „chlupatý“ pavouk, známý z filmových hororů. Ve skutečnosti je to plachý tvor, jehož kousnutí je relativně neškodné (Williams a Goette, 1997). Foelix (1982, viz str. 45 – 46) píše o tarantuli, že její kousnutí není prakticky horší než vosí žihadlo.
„Nejdávnější“ známí pavouci měli „téměř kompletní snovací bradavku, jejíž struktura je docela vyspělá“ (Preston – Mafham, 1991, viz str. 13). Tuhý exoskeleton mnoha pavoukovců napomáhá jak jejich zachování tak rozpoznávání. Mnoho pavouků jsme nalezli velmi dobře zachované coby fosilie (v jantaru a jinde), a jsou snadno rozpoznatelní. Jantar zachoval dokonce i ty členovce, již tuhé krunýře nemají. Fosilní pavouci byli identifikováni z téměř 60 různých čeledí v jediném typu jantaru, a jsou datováni zpět do terciéru a někteří dokonce i z tak dávné doby jako je svrchní křída či ještě dříve (Penney, 2001, 2002; Schawaller, 1983; Poinar, 2000).
Jen v samotné Dominikánské republice bylo v miocenním jantaru rozpoznáno přes 200 druhů pavouků (Penney, 2001). Mnohé z fosilií v jantaru, jež „se datují tak dávno jako 55 miliónů let zpět, patří k rodům, jež stále ještě existují“ (Preston – Mafham, 1996, str. 9). Tihle pavoukovci v jantaru se jeví velmi moderními ve všech ohledech, a dokonce mají i plně rozvinutá vlákna svých sítí (Poinar a Poinar, 1999). V jantaru jsme nalezli i plně moderně vypadající štíry (Poinar a Poinar, 1999, viz str. 76 – 78). Leeming konstatoval, že štíři existují relativně nezměněni od svého prvního výskytu ve fosilních nálezech – tedy před asi 450 milióny let (2004).
Současné fosilní objevy datují moderní formy pavoukovců zhruba 414 miliónů let zpět, „což nutí vědce, aby přehodnotili své názory ohledně…jednoho z nejdůležitějších kroků v evoluční historii“ (Monastersky, 2003, str. 292). Existující exempláře zachované v jantaru podporují závěr, že neexistuje důkaz pro makroevoluci (Shultz, 1994). Běžný názor vědců je ten, že se pavoukovci vyvinuli z nějakého ostrorepovitého zvířete. Jak však připouští Preston – Mafham, „můžeme jenom hádat, jak asi vypadali předkové pavouků a štírů. Ačkoli ještě nemáme definitivní důkaz, věří se, že všichni pavoukovci vyšli ze společného předka“ (Preston – Mafham, 1991, str. 12, 15). Absence předkovských spojovacích článků se týká též štírů a dalších pavoukovců (Poinar et al., 1998; Fortey a Thomas, 1997).
Neodarwinismus musí též vysvětlit evoluci vedlejších orgánů pavoukovců. Evoluce vláknových žláz a snovacích bradavek pavouka nutných pro tvorbu sítí je například vysvětlována domněnkou, že před 180 milióny let bylo pavoučí vlákno prostě jen „výměškem, který se táhl za ním, jak pavouk běžel“ (Kaston, 1966, str. 27). Z tohoto stadia se vlákno vyvinulo ve vlečné lano, dále v lano k cestování, a nakonec vytvořilo pavoučí síť. Problémy s touto myšlenkou zahrnují potřebu vysvětlit, jak pavouk přežíval do doby, než byla snovací bradavka zcela vyvinuta, a vysvětlit fakt, že bradavka produkující vlákno je nezjednodušitelně komplexní a je umístěna jinde než řitní otvor.
Jinou teorii o evoluci pavoučí sítě představuje názor, že původní pavouci užívali alfa keratin (složku pavoučích vláken) k zakrývání svých vajíček (Vollrath, 1992). A když už se jednou vláknové žlázy vyvinuly, musel se též vyvinout program v mozku, jenž řídil tvorbu sítě – vlákno je k ničemu pro chytání potravy, pokud nemá pavouk mentální schopnost upříst síť (Williams, 1992, viz str. 88- 89). Dále, „primitivní sítě nejsou nutně strukturálně jednodušší či méně komplexní než sítě složité“ (Williams, 1988, str. 123).
Pavoučí vlákno produkují snovací (vláknové) žlázy, jež se otvírají z čepů umístěných na snovacích bradavkách (Donovan, 1994). Existuje mnoho druhů vlášení (vláken) – jen taková samička křižáka obecného umí vytvořit nejméně sedm různých druhů vlášení (Vollrath, 1992). Vlášení je bílkovinné povahy a může se dobře zachovat ve fosilních nálezech – pavoučí vlákna byla nalezena už ze středního terciéru, a jedna pavoučí síť se datuje z eocénu (Codington, 1992). Coby fosilie nalézáme i snovací bradavky – po celou historii života zpět až do devonu a karbonu (Donovan, 1994). Nálezy svědčí o tom, že nejstarší známá vlákna a bradavky byly zcela moderní, a neexistují důkazy evoluce snovací bradavky – teorie, že se vyvinula z páru nohou, nenalézá důkazy ve fosilních nálezech (Vollrath, 1992, viz str. 72).
Myriapoda
Podkmeny myriapodních zahrnují stonožky a mnohonožky. Nálezy myriapod existují z celé doby existence života až zpět ke kambriu (Budd, 2001; Kraus a Kraus, 1994). Tato zvířátka mají mnoho párů nohou, buď jeden či dva páry připadají na jeden článek těla (Williams et al., 1991). Mnohonožky jsou zachovány v jantaru a dokonce i v horninách jako výsledek takových procesů jako je potahování, vyztužování a nasycování jejich chitinového exoskeletonu uhličitanem vápenatým.
Dokonce i „jemné struktury jako jsou nožky, anténky, pářící kladélka /gonopody/ a oči“ – všechno bylo krásně zachováno (Donovan a Veltkamp, 1994, str. 355). Výborně zachované fosilie ze spodního devonu, jež odhalují jemné detaily exoskeletonů, byly nalezeny v oblastech izolovaných od hlavních zemských ker jako je Austrálie ((Edgecombe, 1998a, viz str. 172 – 174). Tyto fosilie svědčí pouze o variacích v rámci Bohem stvořených základních typů (baramin), nikoli o evoluci; mnoho z nich je totožných s myriapody existujícími dnes. Myriapoda jsou proto jednou z nejproblematičtějších skupin, v nichž bychom chtěli použít morfologii jako základ pro vytváření fylogenetických rodokmenů (Giribet a Ribera, 2000).
Genetická srovnávání
Problémy s využíváním fosilních nálezů k vystopování původu členovců jsou tak velké, že někteří badatelé zkusili ignorovat zcela fosilie a zvolili jiné metody k vytyčení evolučních rodokmenů, třeba data ze sekvencí DNA. Jelikož jsou tato data DNA často sporná, jiní vědci (jako třeba Meglitsch a Schram, 1991) pominuli jak biochemická, tak DNA data v pokusech o sestavení evolučních rodokmenů. Spolehli se pouze na morfologii žijících zvířat. Je těžké v tomto raném stadiu činit jiné než jen předběžné závěry o užitečnosti genetických srovnávání v konstruování fylogenetických rodokmenů, protože byl prozkoumán jen zlomek členovců. Také nyní poznáváme, že nevíme tolik, jak jsme si mysleli před pár lety, o tom, jak nukleové kyseliny ovlivňují finální fenotypy.
Existující biochemická srovnání nejsou pro neodarwinisty příliš povzbudivá. Burmester (2002) si všiml, že evoluce hemocyaninu je absolutně odlišná v každé ze tří skupin členovců: u klepítkatců, myriapod i korýšů. Jedním z řešení je přesunout původ kmene do ranějších fází vývoje života a předpokládat období „zrychlené“ molekulární evoluce (Vermeij, 1996, str. 525). Toto řešení však vytváří další nesoulady a problémy s fosilními nálezy (Schram a Koenemann, 2001).
Hemocyanin = bezbarvý respiratorní pigment obsahující měď v krevní plasmě nižších mořských živočichů (pozn. překl.)
Vědci mají za to, že nejvhodnějším nukleotidem pro studium evoluce vícebuněčných živočichů je rRNA (Valentine et al., 1996). Při studiu sekvencí ribozomální DNA zjistili Giribet a Ribera (2000) vysokou hladinu sekvenční heterogenity v genu rRNA 18s, že to tak ztížilo práci na fylogenetických srovnáních, že někteří pracovníci mají za to, že tento přístup má jen malý či žádný význam.
Rozdíly mezi daty z molekulární biologie, paleontologie a neontologie (zachování mladších rysů v dospělém stadiu vývoje) zapříčinily to, co Giribet a Ribera (2000) popisují jako zuřivé debaty (viz str. 204). Zmínění vědci uzavírají své výzkumy tak, že data ze sekvencí ribozomální DNA sama o sobě „asi neobsahují dosti informací na to, aby přinesla uspokojivé vysvětlení dlouhé a komplikované evoluční historie členovců“ (str. 225).
Diskuse
Všechny fosilní nálezy členovců dokazují skutečnost, kterou Eldredge zjistil již u trilobitů: že totiž „stabilita byla normou“ , ačkoli se občas skutečně vyskytují „malé“ změny – mikroevoluce (Eldredge, 2000, str. 84). Eldredge také konstatoval, že uplynulé století studia fosilií paleontology neustále přinášelo důkazy o „setrvalosti stabilních druhů po milióny let“ (Eldredge, 2000, str. 85). Po zjištění, že „paleontologie nepřispěla dosud skoro vůbec k evoluční teorii“, navrhl Eldredge (1977, str. 306) několik důvodů, proč tomu tak je. Hlavním důvodem je, že „typický průběh evoluce je tak rychlý, že ji zřídkakdy zachytíme v procesu, v proudu, když prohledáváme fosilní nálezy, abychom se dověděli, jak probíhala (2000, str. 85).
Tento závěr je založen na pouhé spekulaci a je příčinou Gouldova (1977) konstatování, že „hlavní otázky paleontologů po historii života nenašly řešení v rámci darwinovského paradigmatu“ (str. 1).
Praxe obcházení problémů s fosilními nálezy používáním jiných kritérií přináší kladogramy založené výhradně na srovnávání morfologických rysů. Zcela tak ignoruje jak biochemická data, tak fosilní nálezy – obé totiž neposkytuje důkazy pro neodarwinismus. Jak konstatoval Willmer (1990, str. 75), přestože se snažíme seč můžeme, „nedosáhli jsme mnoho porozumění“ evoluci bezobratlých ze studia fosilních nálezů. Běžné jsou na tomto poli nové závěry rušící existující pevně zavedené představy (Cowen, 2003; Doyle, 1996).
Ve fosilních nálezech některých členovců se prokázaly dokonce i moderní symbiotické vztahy. Forézie je symbiotický vztah, zejména mezi členovci a některými rybami, v němž jeden organizmus přenáší jiný organizmus rozdílného druhu. Poinar et al. (1998) zjistili, že mezi štírky a mnoha členovci má forézie dlouhou tradici a je běžná v mnoha případech, jak „demonstruje její setrvalost po milióny let, jak nám dokazují fosilní nálezy“ (str. 78).
Závěry
Kdyby bylo k evoluci došlo, oplývaly by fosilní nálezy (enormně velké) jasnými důkazy pro její podporu. Náš malý přehled potvrdil pozorování specialisty na bezobratlé Roberta Barnese (1980), že „fosilní nálezy nám neříkají téměř nic o evolučních počátcích kmenů a tříd. Spojovací články (přechodné formy) prostě neexistují“ (str. 365). Barnes (1980) též poznamenal, že jedinou výjimkou z této generalizace je zvíře, „jež může představovat předka hlavonožců“ (str. 365). Dodal, optimisticky, že potřebné přechodné formy existují, ale ještě jsme je neobjevili či nerozeznali a další práce je odhalí (Barnes, 1980).
Po dalších dvou dekádách je Barnesův závěr stále platný: potřebné přechodné formy nebyly nalezeny. Fosilní nálezy nám dokumentují, že počet druhů mořských bezobratlých je zhruba konstantní už více než 600 miliónů let evolučního „času“ (Raup, 1976, 1991; Raup a Boyajian, 1988). Problém je prostě ten, že uvažované rodokmeny byly založeny na schématech sestavených podle domněnek založených na evoluční teorii, a tato schémata byla zase užívána jako důkazy teorie použité k jejich sestavení, což je začarovaný kruh (Easton, 1960, viz str. 34).
Ačkoli fosilní nálezy „by měly být rozhodujícím činitelem při výběru z protichůdných názorů na hlavní problémy fylogeneze“ (Willmer, 1990, str. 52), jsou fosilie ignorovány a jakákoli jen trochu schůdná metoda je uplatňována k podpoře teorie všeobecného vývoje. Thomas (2003) připustil, že členovci vládnou světu (alespoň mezi vícebuněčnými zvířaty) a že
bychom rádi věřili, že rozumíme hlavním rysům jejich evolučních vztahů. Bohužel, není tomu tak. Když se naposledy sešli experti na všechny větve členovců, aby diskutovali o jejich fylogenezi, kdysi, roku 1996, nápadně jsme postrádali všeobecnou shodu. Od té doby lze s klidem konstatovat, že prakticky všichni přijímají názor, že členovci jsou monofyletická skupina – to znamená, že jejich společným předkem bylo cosi, co bychom určili jako členovce. Co je nad tento dosti skromný bod, v němž všichni spolu souhlasí, je opředeno vášnivými diskusemi – třeba evoluční vztahy v rámci (uvnitř) a mezi čtyřmi hlavními existujícími skupinami členovců… Jedním z mála bodů, na němž se všichni shodli, je fakt, že hexapoda jsou monofyletická, to znamená, že se vyvinuli pouze jednou z jediného společného šestinohého předka. (str. 1854).
Thomas konstatoval, že i tato shoda ohledně šestinohých tvorů byla nyní otřesena výzkumy DNA (viz Nardi et al., 2004). Další práce učinila darwinistický světový názor ještě nejasnějším. Jak konstatoval Valentine (1989), „spekulace týkající se evolučních vztahů mezi kmeny žijících živočichů začaly už krátce po zveřejnění Darwinových původních prací a pokračují dodnes“ (str. 2272). Toto konstatování platí přesto, že dnešní fosilní nálezy „se prostírají o sto miliónů let dále v čase než nálezy známé v Darwinově době“ (Valentine, 1989, str. 2272). Současná situace, jak ji sumarizuje Aguinaldo (1997), potvrzuje moje hodnocení:
„Členovci jsou nejrozmanitější skupinou zvířat, ale navzdory bohatým fosilním nálezům členovců a století studia zůstávají jejich fylogenetické vztahy nejasné. Taxony dříve považované za sesterské skupiny členovců zahrnují kroužkovce, drápkovce, želvušky a jiné, ale hypotézy fylogenetických vztahů si odporují“ (str. 489).
Kladogram je rozvětvený diagram různých forem života umístěných do rodokmenu podle srovnávání vybraných rysů, jež se, jak se předpokládá, odvíjejí od společného předka. Po prostudování 179 publikovaných kladogramů konstatoval Wills (2001), že „v jistých ukazatelích jsou kladogramy členovců méně v souladu s fosilními nálezy než rozsáhlý vzorek kladogramů různých jiných taxonů“ (str. 201). Důvodem této nízké míry shody jsou prý (jak se často tvrdí) mezery ve fosilních nálezech. Avšak fosilie členovců, jež jsou nejtrvanlivější, jako jsou trilobiti, vykazují jednu z nejmenších shod s kladogramy, a „pro některé rodokmeny téměř jakákoli namátkou vybraná řada či namátková topologie rodokmenu vykazuje shodu větší“ (Wills, 2001, str. 206; viz též Weygoldt, 1996). Jinými slovy, výběr rozdílných rysů pro srovnávání vede k rozdílným kladogramům.
Všechny tyhle problémy podporují názor, že mnoho základních typů členovců (baramin – základní stvořený typ) bylo stvořeno zvlášť. Makroevoluce opět vykazuje rysy méně spolehlivé než je koncept speciálního Stvoření.
Poděkování
Chtěl bych poděkovat Waynu Frairovi Ph.D., Bertu Thompsonovi Ph. D., Cliffordu Lillovi M. A., Georgu Howemu Ph. D., Johnu Woodmorappovi M. A. a Jodymu Allenovi za jejich připomínky k první verzi tohoto článku. Děkuji také Emmettu L. Williamsovi Ph. D. za pomoc při vyhledávání starších referátů o původu členovců Stvořením a jednomu anonymnímu expertu za několik podnětných připomínek.
Odkazy
CRSQ: Creation Research Society Quarterly
Aguinaldo, A.M.A., J.M. Turbeville, L.S. Linford, M.C. Rivera, J.R. Garey R.A. Raff, and J.A. Lake. 1997. Evidence for a clade of nematodes, arthropods and other moulting animals. Nature 387:489–493.
Alonso, J., A. Arillo, E. Barron, J.C. Corral, J. Grimalt, J.F. Lopez, R. Lopez, X. Martinez-Delclos, V. Ortuno, E. Penalver, and P.R. Trincao. 2000. A new fossil resin with biological inclusions in Lower Cretaceous deposits from Alava (northern Spain, Basque-Cantabrian Basin). Journal of Paleontology 74(1):158–178.
Bergman, J. 2004. Insect evolution: a major problem for Darwinism. Creation ex Nihilo Technical Journal 18(2):91–97.
Delclos, V. Ortuno, E. Penalver, and P.R. Trincao. 2000. A new fossil resin with biological inclusions in lower cretaceous deposits from alava (northern Spain, Basque-Cantabrian Basin). Journal of Paleontology 74:158–178.
Anderson, D.T. (editor). 1998. Invertebrate Zoology. Oxford University Press, South Melbourne, Australia.
Barnes, R.D. 1980. Invertebrate beginnings. Paleobiology 6:365–370.
Barnes, R.D. 1987. Invertebrate Zoology (5th Edition). Saunders, Philadelphia, PA.
Budd, G.E. 1997. Stem group arthropods from the Lower Cambrian Sirius Passet fauna of North Greenland. In R.A. Fortey and R.H. Thomas (editors), Arthropod Relationships, pp. 125–136. Chapman and Hall, London, UK.
Budd, G.E. 2001. A myriapod-like arthropod from the Upper Cambrian of East Siberia. Palaeontologische Zeitschrift 75:37–41.
Burmester, T. 2001. Molecular evolution of the arthropod hemocyanin superfamily. Molecular Biological Evolution 18:184–195.
Burmester, T. 2002. Origin and evolution of arthropod hemocyanins and related proteins. Journal of Comparative Physiology B: Biochemical, Systemic, and Environmental Physiology 172:95–107.
Chaloner, W.G., A.C. Scott, and J. Stephenson. 1991. Fossil evidence for plant-arthropod interactions in the Palaeozoic and Mesozoic. Philosophical Transactions: Biological Sciences 333:177–185.
Clarkson, E.N.K. 1986. Invertebrate Palaeontology and Evolution (2nd Edition). Unwin Hyman, London, UK.
Clarkson, E.N.K. 1998. Invertebrate Palaeontology and Evolution (3rd Edition). Unwin Hyman, London, UK.
Codington, L.A. 1992. Fossil spider web from the Eocene of Western Colorado. Geological Society of America Abstracts with Program 24:A344.
Cook, M.A. 1968. William J. Meister discovery of human footprint with trilobites in a Cambrian formation of western Utah. CRSQ 5:97.
Cowen, R. 2003. Genetic trickery probes tropical parasites. Science News 138:292.
Davis, L.J. 1968. Rockhound finds puzzling fossil. CRSQ 5:102.
DeYoung, D. 2002. Vision. CRSQ 38:190–192.
Donovan, S.K. 1994. Insects and other arthropods as trace-makers in non-marine environments and palaeoenvironments. In S.K. Donovan (editor), The Palaeobiology of Trace Fossils, pp. 200–220. The Johns Hopkins University Press, Baltimore, MD.
Donovan, S.K., and C.J. Veltkamp. 1994. Unusual preservation of late Quaternary millipedes from Jamaica. Lethaia 27:355–362.
Doyle, P. 1996. Understanding Fossils: An Introduction to Invertebrate Paleontology. Wiley, New York, NY.
Easton, W.H. 1960. Invertebrate Paleontology. Harper, New York, NY.
Edgecombe, G.D. (editor). 1998a. Arthropod Fossils and Phylogeny. Columbia University Press, New York, NY.
Edgecombe, G.D. 1998b. Devonian terrestrial arthropods from Gondwana. Nature 394:172–175.
Eldredge, N. 1977. Trilobites and evolutionary patterns. In A. Hallam (editor), Patterns of Evolution, pp. 305–332. Elsevier, Amsterdam, The Netherlands.
Eldredge, N. 2000. The Triumph of Evolution and the Failure of Creationism. W.H. Freeman, New York, New York.
Foelix, R. 1982. Biology of Spiders. Harvard University Press, Cambridge, MA.
Fortey, R.A., and R.H. Thomas (editors). 1997. Anthropod Relationships. Chapman and Hall, London.
Gayrard-Valy, Y. 1994. Fossils: Evidence of Vanished Worlds. Abrams, New York, NY.
Giribet, G., and C. Ribera. 2000. A review of arthropod phylogeny: New data based on ribosomal DNA sequences and direct character optimization. Cladistics 16:204–231.
Gould, S.J. 1977. Eternal metaphors of palaeontology. In A. Hallam (editor), Patterns of Evolution, pp. 1–26. Amsterdam: Elsevier, Amsterdam, The Netherlands.
Hallam, A. 1977. Patterns of Evolution as Illustrated by the Fossil Record. Elsevier Scientific, Oxford, New York, NY.
Hamilton, H.S. 1986. The jumping spider’s wondrous eyes. CRSQ 23:63–64.
Harper, D.A.T., and M.J. Benton (editors). 2001. History of biodiversity. Geological Journal 36:187–210.
Helder, M. 1997. The extra special courtship of horseshoe crabs. Creation Matters 2(1):1–3.
Kaston, B.J. 1966. Evolution of the web. Natural History 75:27–32.
Kraus, O., and M. Kraus. 1994. Phylogenetic system of the tracheata (mandibulata): on “myriapoda”: insecta interrelationships, phylogenetic age and primary ecological niches, Verhandlungen des Naturwissenschaftlichen Vereins in Hamburg 34:5–31.
Kaufmann, D.A. 1977. Phylogenetic development of adipose tissue in animals. CRSQ 13:214–215.
Lammerts, W.E. 1974. Insect family tree may become a forest. CRSQ 11:124–125.
Le Conte, Joseph. 1908. Elements of Geology. D. Appleton and Company, New York, NY.
Leeming, J. 2004. Book review: scorpions of southern Africa. Wildwatch—African Wildlife and Conservation. http://www.wildwatch.com/resources/reviews/scorpionsSA.asp (as of December, 2005).
Levi-Setti, R. 1993. Trilobites. The University of Chicago Press, Chicago, IL.
Manning, A. 2003. The Wright brothers’ airplane compared to insect flight design. CRSQ 40:1–7.
Manton, S.M. 1973. Throw Away Your Zoology Textbooks. Queen Mary College, London, UK.
Margulis, L., and D. Sagan. 2002. Acquiring Genomes: A Theory of the Origins of Species. Basic Books, New York, NY.
Meglitsch, P.A. 1967. Invertebrate Zoology. Oxford University Press, New York, NY.
Meglitsch, P.A. and F.R. Schram. 1991. Zoology (3rd Edition). Oxford University Press, New York, NY.
Meister, W.J., Sr. 1968. Discovery of trilobite fossils in shod footprint of human in “trilobite beds”— a Cambrian formation, Antelope Springs, Utah. CRSQ 5(3): 97–101.
Monastersky, R. 2003. Fossils push back origin of land animals. Science News 138:292.
Morris, S.C. 2000. The Cambrian “explosion”: slow-fuse or megatonnage? Proceedings of the National Academy of Science 97:4426–4429.
Nardi, F., G. Spinsanti, J.L. Boore, A. Carapelli, R. Dallai, and F. Frati. 2003. Hexapod origins: monophyletic or paraphyletic? Science 299:1887–1889.
Penney, D. 2001. Advances in the taxonomy of spiders in Miocene Amber from the Dominican Republic (Arthropoda: Araneae). Palaeontology 44:987–1009.
Penney, D. 2002. Spiders in Upper Cretaceous amber from New Jersey (Arthropoda: Araneae). Palaeontology 45:709–724.
Poinar, G.O., Jr. 2000. Heydenius araneus n.sp. (Nematoda: Mermithidae), a parasite of a fossil spider, with an examination of helminths from extant spiders (arachnida: Araneae). Invertebrate Biology 119:388–393.
Poinar, G.O., B.P.M. Curcic, and J.C. Cokendolpher. 1998. Arthropod phoresy involving pseudoscorpions in the past and present. Acta Arachnologica 47:79–96.
Poinar, G.O., and R. Poinar. 1999. The Amber Forest. Princeton University Press, Princeton, NJ.
Preston-Mafham, R.. 1991. The Book of Spiders. Chartwell Books, Edison, NJ.
Preston-Mafham, R., and K. Preston-Mafham. 1996. The Natural History of Spiders. The Crowood Press, Ramsbury, Marlborough Wiltshire, UK.
Prokop, R. 1995. Fossils. Magna, Leicester, UK.
Raup, D.M. 1976. Species diversity in the phanerozoic: an interpretation. Paleobiology 2:289–297.
Raup, D.M. 1991. A kill curve for Phanerozoic marine species. Paleobiology 17:37–48.
Raup, D.M., and G.E. Boyajian. 1988. Pat terns of generic extinction in the fossil record. Paleobiology 14:109–125. Ruppert, E.E., R.S. Fox, and R.D. Barnes. 2004. Invertebrate Zoology: a Functional Evolutionary Approach (7th Edition). Brooks/Cole, Belmont, CA. Schawaller, W. 1983. The spider family Her siliidae in Dominican amber Stuttgart amber collection Arachnida araneae. Stuttgarter Beitraege zur Naturkunde Serie B (Geologie und Palaeontologie) 79:1–10. Schram, F.R., and S. Koenemann. 2001. Developmental genetics and arthropod evolution: part I, on legs. Evolution and Development 3:343–354. Shear, W.A., P.A. Gensel, and A.J. Jeram. 1996. Fossils of large terrestrial ar thropods from the Lower Devonian of Canada. Nature 384:555–557. Sherwin, F., and M. Armitage. 2003. Trilobites—the eyes have it! CRSQ 40:172–174. Shultz, J.W. 1994. The limits of stratigraphic evidence in assessing phylogenetic hy potheses of recent arachnids. Journal of Arachnology 22:169–172. Thomas, R.H. 2003. Wingless insects and plucked chickens. Science 299:1854– 1855. Tootill, E. 1988. The Facts on File Diction ary of Biology. Revised and Expanded Edition. Facts on File Publications, New York, NY. Twenhofel, W.H., and R.R. Shrock. 1935. Invertebrate Paleontology. McGraw Hill, New York, NY. Valentine, J.W. 1989. Bilaterians of the Precambrian-Cambrian transition and the annelid-arthropod relationship. Proceedings of the National Academy of Science 86:2272–2275. Valentine, J.W., D.H. Erwin, and D. Jablon ski. 1996. Developmental evolution of metazoan bodyplans: the fossil evidence. Developmental Biology 173:373–381. Vermeij, G. 1996. Animal origins. Science 274:525. Vollrath, F. 1992. Spider webs and silks. Scientifi c American 266:70–76. Weygoldt, P. 1996. Evolutionary morphology of whip spiders: towards a phylogenetic system. Journal of Zoological Systematics and Evolutionary Research 34:185–202. Whittington, H.B. 1992. Trilobites. The Boydell Press, Rochester, NY. Williams, E.L. 1988. Panorama of science. CRSQ 25:123–124. Williams, E.L. 1992. Spiders as engineers. CRSQ 28:88–89. Williams, E.L, and R.L. Goette. 1997. Ta rantula goes acourtin’, and he does roam. CRSQ 34:3–4. Williams, E.L, G.F. Howe, and R.R. White. 1991. A desert millipede: evolution or design?—an introduction. CRSQ 28:7–16. Willmer, P. 1990. Invertebrate Relationships. Cambridge University Press, Cambridge, MA. Wills, M.A. 2001. How good is the fossil re cord of arthropods? an assessment using the stratigraphic congruence of clado grams. Geological Journal 36:187–210. Wills, M.A, D.E.G. Briggs, and R.A. Fortey. 1997. Evolutionary correlates of ar thropod tagmosis: scrambled legs. In R.A. Fortey and R.H. Thomas (editors), Arthropod Relationships, pp. 57–65. Chapman and Hall, London, UK.