Z creation.com/multicellularity přeložil Jakob Haver – 10/2019. Translation granted by Creation.com – Přeloženo s povolením od Creation.com.
Celá evoluce předpokládá buď vylepšení nějakého předcházejícího systému, který se přizpůsobí novým potřebám, nebo přenesení vnějšího genu, který přinese nové informace se stejným cílem jako v prvním případě. Předpokládá se, že i takové systémy, které zjevně vyžadují zcela nové struktury (např. peří), byly upravené z původně existujících struktur. V historii evoluce jsou však dvě události, kdy toho bylo potřeba mnohem víc – vznik života a vznik koordinované mnohobuněčnosti.
Co je nezbytné pro vývoj mnohobuněčnosti
Stejná genetická informace
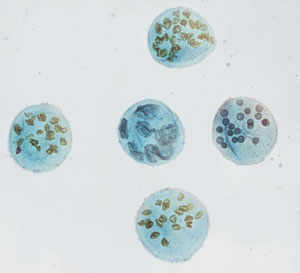
Valeč koulivý (Volvox) nesplňuje požadavky k dosažení skutečné mnohobuněčnosti. Foto Karl Dolenc, iStock.com.
Prvním požadavkem pro vznik mnohobuněčnosti je totožnost genetické informace u všech zúčastněných buněk. Dobrý přehled toho, proč je k životaschopnosti samostatného mnohobuněčného organismu potřebná genetická identita, poskytují Wolpert a Szathmáry:
„První krok ve vývoji složitého organismu je ustanovení vzorce buněk s různými stavy, které se mohou lišit podle svých drah … Procesy modelování vyžadují mezi buňkami a uvnitř buňky signalizaci, což nakonec vede k zapnutí či vypnutí genu. Takový proces může vést ke spolehlivým vzorcům buněčné aktivity jen tehdy, mají-li všechny buňky stejnou sestavu genů a řídí se stejnými pravidly [přidáno zdůraznění].”1
Nevychází-li jejich činnost ze stejného genetického plánu, neexistuje žádná záruka, že buňky budou schopny správně komunikovat při své koordinované činnosti.
Nová úroveň biologické organizace
Evoluce potřebuje pro rozvoj koordinované činnosti víc než jen pouhé zvětšení existujícího systému; je nutné vytvoření ex nihilo zcela nového systému vhodně uspořádaných koordinovaných buněk tak, aby vytvořily samostatný mnohobuněčný organismus. S tím souhlasí i Nedelcu a Michod:
„Současné hierarchické uspořádání života odráží ve vývoji celků řadu přechodů, např. z genů na chromozómy, z prokaryotických buněk na eukaryotické, z jednobuněčných na mnohobuněčné celky, z mnohobuněčných organismů na kmeny. Během těchto vývojových přechodů vznikají nové úrovně biologického uspořádání [přidáno zdůraznění]”.2
Williams mluví o neredukovatelné struktuře buňky a jako všeobecný příklad uvádí autopoézu (sebe-produkce).3 Popisuje pět organizačních úrovní, nutných pro vznik autopoézy ve všem živém:
- Dokonale čistá biochemie, specifická pro každou jednotlivou molekulu.
- Molekuly s vysoce specifickými strukturami.
- Vysoce strukturované, funkčně integrované molekuly.
- Komplexně regulované, informacemi řízené metabolické procesy.
- Inverzně kauzální, meta-informační (informace o informaci) strategie pro přežití jednotlivce a druhu.
Kromě toho, každá úroveň je větší než jen prostý součet úrovní, které ji tvoří, takže jediný způsob, jak tyto úrovně vysvětlit je pomocí informací.
„Všechny úrovně stojí jedna na druhé, ale jednotlivě je nelze vysvětlit z hlediska té úrovně pod ní. A mezi základní úrovní (což je dokonale čistá kompozice) a přírodním prostředím je nepřekročitelná propast.“4
K Williamsově hierarchii autopoézy bych rád přidal další úroveň struktury, která se nalézá pouze u mnohobuněčných organismů: mezibuněčná koordinace. Organismus má pro přežití a reprodukci své strategie na uspořádání a diferenciaci svých buněk. K tomu je potřebná komunikační síť mezi buňkami, která řídí umístění a četnost každého typu buňky ve prospěch celého organismu. Zásadní součástí této organizace je buněčná diferenciace, která je u mnohobuněčných organismů všudypřítomná. Tuto úroveň nelze vysvětlit pouhým součtem částí a buněk, je také nutná koordinace jednotlivých buněk z organizační úrovně nad nimi.
Biolog Eric Davidson5 rozlišuje v mnohobuněčných organismech čtyř-úrovňovou hierarchii řízení, která tvoří genovou regulační síť. Tato genová regulační síť je pro vývoj od jednobuněčné zygoty k plnohodnotnému mnohobuněčnému organismu nezbytná. Řečeno v rámci přibližné Linnéovy klasifikace, hierarchie se skládá ze základních jader6, která zhruba vymezují tělesný plán kmene; dále doplňkových modulů7 a vstupních/výstupních vazeb8, které přibližně určují tělesnou strukturu třídy, řádu a rodu, a baterie diferenciačních genů9, které provádějí závěrečné fáze vývoje a podílejí se na variacích na úrovni rodů a druhů.
Strategie oprav a údržby
Nedílnou součástí k přežití dospělého mnohobuněčného jedince jsou strategie oprav a údržby, protože k výběru těch reprodukčně nejprůbojnějších buněk pracuje buněčný výběr s buněčnými populacemi a mnohobuněčnými organismy. To musí být řízeno na úrovni organismů, aby byla zachována tělesná integrita. Aby to bylo možné, podléhá většina systémů mnohobuněčných tvorů procesu postupné diferenciace.10 V tomto systému jsou nezbytné multipotentní11 kmenové buňky, i když jsou udržovány na nízké úrovni populace.
Buněčný výběr vs. integrita organismu12
Je jasné, že pokud se mnohobuněčnost měla vyvinout, čelí evoluce těžkému rozporu: buněčný výběr vs. integrita organismu. Výběr bude na úrovni jednotlivých buněk upřednostňovat takové buňky, které se lépe množí. Pokud se však těmto buňkám umožní nekontrolovatelně se množit v mnohobuněčném organismu, neúprosně zničí integritu organismu, organismus poškodí nebo usmrtí a přivodí smrt také „odolnějším“ buňkám.13
Na úrovni organismů bude výběr upřednostňovat vlastnosti, které zachovávají integritu organismu a snaží se řídit reprodukci buněk i nad rámec toho, co je potřeba. S tím souhlasí Pepper a kol.:
„Mnohobuněčné organismy se nemohly objevit jako funkční prvky dříve, než výběr na úrovni organismu dospěl k vývoji mechanismů, potlačujících výběr na buněčné úrovni.“14
To však přivádí evolucionisty k záhadě: jak se vyvinuly mnohobuněčné organismy z jednobuněčných, jestliže buněčný výběr a výběr na úrovni organismu jdou navzájem vysloveně proti sobě? Mnohobuněčný organismus se snaží řídit reprodukci podle toho, co je potřebné na vyšší organizační úrovni; samostatná buňka se snaží větší reprodukcí překonat své konkurenty.
Vypadá to, že pro mnohobuněčnost jsou nezbytné mechanismy apoptózy (programovaná buněčná smrt), kdy některé buňky dostanou pokyn zemřít během vývoje nebo proto, že jsou porušené. Takové mechanismy jsou nepředstavitelně složité a nepochybně neredukovatelně komplexní.15 Vysvětlení existence takového mechanismu bez inteligentního designu se dá rovnou nazvat přehlídkou marnosti.16
Spolupráce a kolonie: na půli cesty?
Předpokládá se, že mnohobuněčnost se vyvinula cestou spolupráce a kolonizace organismů. U jednobuněčných organismů taková spolupráce nastává. Například Salmonella typhimurium se při invazi umí uspořádat do dvou linií – první řada se vrhá k sebevražednému útoku a druhá řada tímto bojovným zmatkem, vyvolaným první vlnou proklouzne.17 Tudíž mezi jednobuněčnými organismy dochází k nějaké komunikaci, která umožňuje spolupráci.
Kolonie si vytváří mnoho organismů. Nicméně ve většině těchto kolonií si jednotlivé buňky zachovávají schopnost kolonii „opustit“, nastanou-li pro to výhodnější podmínky. Koloniální systémy sice spolupracují, ale nemají žádný regulační systém, který by přinutil buňky spojit se jako plnohodnotný samostatný celek výběru. Kromě toho kolonie organismů se může rozpadnout, aniž by jednotlivé buňky utrpěly nějakou zvláštní škodu, na rozdíl od mnohobuněčného organismu, který kdyby se rozpadl, vážně ho to poškodí nebo zahyne. S tím souhlasí i Michod a kol.:
„Takové kolonie a skupiny mohou přetrvávat a měnit se podle různých možností, v závislosti na vlastnostech skupiny a jejím konkrétním složení. Zpočátku je zdatnost skupiny průměrem zdatností jednotlivců nižší úrovně, ale jak dochází k vývojovým přechodům, skupinová zdatnost se od zdatnosti složek nižších úrovní odděluje. Podstatou evolučního přechodu v samostatnou individualitu je, že jednotlivci nižší úrovně se musí „vzdát“ svého „práva“ na zdatnost, tj. mohou vzkvétat a množit se pouze ve prospěch nové jednotky vyšší úrovně.“18
 Některé bakteriální kolonie se však jeví jako by byly svázány a vykazují určitou specializaci, například někteří zástupci rodu Volvolaceae, jako je Volvox carteri. Důvodem, proč bakteriální kolonie nesplňují parametry skutečných mnohobuněčných organismů je to, že nemají totipotentní dělení19 a „nesmrtelnosti“:20
Některé bakteriální kolonie se však jeví jako by byly svázány a vykazují určitou specializaci, například někteří zástupci rodu Volvolaceae, jako je Volvox carteri. Důvodem, proč bakteriální kolonie nesplňují parametry skutečných mnohobuněčných organismů je to, že nemají totipotentní dělení19 a „nesmrtelnosti“:20
Vypnutí nesmrtelnosti a totipotence se u V. carteri ukázalo jako nemožné: tyto znaky jsou vyjádřeny buď společně a plně (např. V. gonidii) nebo vůbec (např. v somatických buňkách). Nesmrtelnost a totipotence jsou tedy u V. carteri stále pevně zapnuty, stejně jako u jejich jednobuněčných předků. K podpoře tohoto názoru přispívá fakt, že u V. carteri chybí mutantní somatické buňky podobné rakovině, ve kterých se znovu získá nesmrtelnost, nikoli však totipotence. Nicméně existují mutantní formy V. carteri…, ve kterých somatické buňky znovu získávají nesmrtelnost i totipotenci, ale v žádné z těchto mutací nejsou vyjádřeny tyto dva rysy částečně nebo odlišně (např. omezená mitotická kapacita nebo multipotence).“21
To znamená, že diferenciace v kolonii se může rozšířit pouze na dva různé typy buněk a dál už ne. Protože řasy volvocine nejsou schopny vypnout totipotenci a nesmrtelnost, nemohou vytvářet nové somatické buňky a v důsledku toho nemohou jako organismus moc dlouho přežít. Jinými slovy, v životní formě řas volvocine nejsou žádné strategie údržby či oprav, takže jim chybí jeden ze klíčových rysů skutečné mnohobuněčnosti.
Příležitosti pro další výzkum
Mou snahou zde bylo předložit některé základní požadavky, které musí být splněny pro vývoj skutečné mnohobuněčnosti. Mezi všemi zúčastněnými buňkami skutečné mnohobuněčnosti musí existovat genetická totožnost. Jako další úroveň v živé organizaci slouží mezibuněčná koordinace, kterou nelze zredukovat jen na součet jejích částí. V regulační architektuře existuje hierarchie na čtyřech úrovních, které jsou nezbytné k pokračování životaschopného plánu vývinu. Opravy a údržba vyžadují jednu nebo více skupin nediferencovaných, obecně multipotentních kmenových buněk. Buněčný výběr a integrita organismu zůstávají diametrálně protichůdné, a to pro evoluci představuje velmi těžký, dosud nepřekonaný problém. Jednobuněčné organismy v koloniích nesplňují podmínky pro mnohobuněčné jedince proto, že tuto čtyř-úrovňovou hierarchii nemají a postrádají mechanismy údržby a oprav organismu.
Toto je opomíjená oblast kreacionistického výzkumu, která skýtá množství příležitostí pro další zkoumání.
Odkazy a poznámky
- , L. and Szathmáry, E., Evolution and the egg, Nature 420:745, 2002. Return to text.
- Nedelcu, A.M. and Michod, R.E., Evolvability, Modularity, and Individuality During the Transition to Multicellularity in Volvocalean Green Algae; in: Schlosser, G. and Wagner, G. (Eds.), Modularity in development and evolution, Univ. Chicago Press, Chicago, IL, pp. 466–489, 2003; p. 466.
- Williams, A., Life’s irreducible structure—Part I: autopoiesis, Journal of Creation 21(2):109–115, 2007; <creation.com/autopoiesis>. Return to text.
- Williams, ref. 3, p. 111.
- Davidson, E.H., The Reguatory Genome: Gene Regulatory Networks in Development and Evolution, Academic Press, Burlington, MA, pp. 195–196, 2006. For the definitions of terms in refs. 6–9, see p. 128.
- Jádra jsou konzervované dílčí okruhy skládající se z regulačních genů, které vzájemně interagují a jsou určeny pro specifické vývojové funkce.
- Moduly plug-in jsou obvyklé přípojné obvody, které se používají pro mnoho různých funkcí při vývinu.
- Vazby vstupu a výstupu jsou řídící regulátory na stejné rovině DNA jako gen, na kterém pracují, a buď je zapínají, nebo vypínají, v závislosti na stupni vývinu.
- baterie diferenciačních genů jsou sady genů, které reagují na běžnou sadu regulátorů buněčného typu, které kódují na úrovni proteinu funkční a strukturální vlastnosti tohoto typu buněk.
- Doyle, S., Serial cell differentiation: intricate system of design, Journal of Creation 22(2):6–8, 2008.
- Multipotentní znamená, že buňka má schopnost diferencovat se do více typů buněk. Není totipotentní, tedy nemůže obnovit celý organismus, ale může obnovit určité tkáně a typy buněk.
- Pro podobnou diskusi týkající se sériové diferenciace, viz Doyle, ref. 10. Michod, R.E., Cooperation and conflict in the evolution of individuality I. Multilevel selection of the organism, The American Naturalist 149:607–645, 1997.
- Pepper, J.W., Sprouffske, K. and Maley, C.C., Animal cell differentiation patterns suppress somatic evolution, PLoS Comput. Biol. 3(12):2532–2545, 2007; p. 2533; <www.ploscompbiol.org/article/info:doi/10.1371/journal.pcbi.0030250>.
- Bell, P., Apoptosis: cell ‘death’ reveals Creation, Journal of Creation (TJ) 16(1):90–102, 2002; <creation.com/apoptosis>.
- Bell, P., The non-evolution of apoptosis, Journal of Creation (TJ) 18(1):86–96, 2004.
- Ackermann, M. et al., Self-destructive cooperation mediated by phenotypic noise, Nature 454:987–990, 21 August 2008.
- Michod, R.E., Nedelcu, A.M. and Roze, D., Cooperation and conflict in the evolution of individuality IV. Conflict mediation and evolvability in Volvox carteri, BioSystems 69:95–114, 2003.
- Zde totipotence znamená schopnost vytvářet nový organismus.
- e. kapacita pro pokračující dělení buněk bez konce.
- Michod, et al., ref. 18, p. 105.