II.
VZNIK PRAŽIVOTA ČIROU NÁHODOU?
A. VZNIK STAVEBNÍHO MATERIÁLU ŽIVOTA
1. Vznik stavebního materiálu z chemického hlediska
Millerovy pokusy (I) i pokusy jiných přírodovědců posílily v odborných kruzích všeobecné přesvědčení, že kyseliny, jež tvoří stavební materiál a bázi života, vznikly čirou náhodou a normálními, neusměrňovanými, neinteligentními chemickými reakcemi. V praatmosféře země, jež se skládala hlavně z vodní páry, amoniaku, metanu, dusíku a kyseliny uhličité, docházelo k elektrickým výbojům nebo bleskům, při nichž se spontánně syntetizovala malá množství různých biologicky potřebných aminokyselin. Miller provedl podobné pokusy v laboratoři a vytěžil přitom malé, ale zřetelné množství stavebního materiálu života (aminokyseliny) (I). Podle mínění odborných kruhů tím vyřešil problém, jak vznikly původní biologicky potřebné aminokyseliny.
Když se nyní takové aminokyseliny za vhodných podmínek rozpustí ve vodě, spojí se po dvou, při čemž se uvolní jedna molekula vody, a vytvoří dipeptid (II)
(I) S. L. MILLER: Science 117 (1953), 528; J. Amer. chem. spol. 77 (1955), 2351.
(I) CH4 + H2O + NH3 + energie R-CH-COOH
NH2
(metan+voda+amoniak (hmota) + energie (např. blesk)
= aminokyseliny (stavební materiál života)
kondenzační reakce
(II) R-CH-COOH + R-CH-COOH ================== R·CH-COOH+H2O
NH2 NH2 NH-CO-CH
NH2
(aminokyselina I - aminokyselina II se sloučí v dipeptid + voda)
Aminová skupina (NH2) aminokyseliny (R-radikál) se sloučí s karboxylovou skupinou (COOH) druhé aminokyseliny a přitom vyloučí HOH (= voda H2O), aby vytvořila dipeptid. Prověřuje-li se vzorec dipeptidu, zjistí se, že má stále ještě jednu skupinu NH2 a jednu skupinu COOH, takže se dipeptid může sloučit s nějakým jiným dipeptidem nebo monopeptidem či nějakou aminokyselinou, aby vytvořil quadrupeptid nebo tripeptid atd. (3). Přitom se opět uvolní jedna molekula vody:
kondenzační
reakce R' R'
(III) R-CH-COOH+R-CH-COOH=========R-CH-CO-NH-CH-CO-NH-CH-COOH+H2O
NH-CO-CH-R' NH-CO-CH-R' NH-CO-CH-R'
NH2 NH2 H2
(dipeptid I + dipeptid II se sloučí v quadripeptid + voda)
Provádí-li se další kondenzace (jak se nazývá tento druh reakce), dojde se nakonec - vytvoří-li se řetězec dlouhý několik tisíc stavebních kamenů aminokyselin - k proteinoidům, jež se trochu podobají bílkovinám živé buňky. Dříve se dokonce tvrdívalo, že sám život jsou jenom kondenzační reakce, při nichž se vylučuje voda. Bílkoviny života se tedy skládají z takových aminokyselin, jenomže tyto řetězce jsou velmi dlouhé. Může v nich být tisíce zbytků aminokyselin.
Protože se aminokyseliny různých druhů mohou samovolně vytvářet pomocí elektrických výbojů v praatmosféře - Miller to dokázal - a protože se aminokyseliny ve vodním roztoku samovolně slučují (podle vzorce II a III) na peptidy, polypeptidy a proteinové substance, je tím podle názoru odborných kruhů položen základ pro spontánní biogenezi (vznik praživota). Stavební materiál života (aminokyseliny) a peptidy (základ bílkovin) se samovolně vytvářejí z hmoty a energie. Tak je mnoho odborníků a většina příslušných učebnic přesvědčena o tom, že z přírodovědeckého hlediska už vůbec nic nestojí v cestě náhodnému, spontánnímu vzniku života. Jestliže tu jednou byla energie a praatmosféra, pak se časem musela objevit (a to se dnes tvrdí) i živá protoplasma a tím i rodokmen života. Neboť podle Alexandra Ivanoviče Oparina se jednoho dne, když bylo v praoceánu dostačující množství životních bílkovin, vytváří mikrosféra nebo koacervát, primitivní buňka, která se začne replikovat, tzn. množit buněčným dělením. Po tomto stadiu se už výběr postará o automatický vzestupný vývoj až k homo sapiens.
Tak vznikl, podle všeobecně uznávaného mínění odborníků, stavební materiál první živé buňky, když v praatmosféře čirou náhodou udeřil blesk. Když už se jednou v praoceánu utvořila kolonie živých buněk, začal mezi nimi závod o živiny a životní prostor - přirozený výběr, jehož výsledkem byl naprosto automatický vzestupný vývoj (evoluce) v neodarwinistickém smyslu, bez jakéhokoliv plánování. Buňky, u nichž došlo k nejpříznivějším mutacím (ovšem čirou náhodou), se prosadily a zanechaly více potomstva než buňky s méně příznivými mutacemi. Podle této teorie, v průběhu mnoha milionů let, musí tento výběr vést až k nejvyššímu biologickému stadiu vývoje (homo sapiens). Člověk tedy čirou náhodou a výběrem musí automaticky vzniknout všude tam, kde jsou dány podmínky (náhoda, energie a výběr). Proto mnozí neodarwinisté tvrdí, že člověk (nebo nějaký rovnocenný druh na srovnatelně vysokém biologickém stupni vývoje) musí časem vzniknout a také vznikne na jiných planetách. Všude ve vesmíru (na jiných planetách), kde jsou splněny čistě fyzikální a chemické předpoklady, dojde k evolučnímu procesu až k vysoce vyvinutému životu. Proto se hledá život - a také život nadaný inteligencí - na jiných planetách a v jiných galaxiích. To je také smysl meziplanetárních raket, které mají experimentálně zjistit, zda je na jiných planetách život. Podle neodarwinistických teorií by se musely nacházet různé druhy života všude tam, kde jsou splněny podmínky pro jeho rozvoj.
Nyní se tedy podíváme na přírodovědeckou oprávněnost těchto všeobecně uznávaných tezí.
2. Spontánní biogeneze v praoceánu?
Mohlo by v praoceánu dojít k biogenezi z aminokyselin a polypeptidů podle vzorců (I-III), jimiž jsme se právě zabývali? Učebnice biologie tvrdí téměř jednohlasně, že tomu tak je. Avšak už jen letmé chemické prověření uvedených rovnic dává na tuto otázku jednoznačně zápornou odpověď.
Kdo prověřuje tyhle rovnice zjistí, že uvedené reakce probíhají obojím směrem, tedy i zpět. Záleží na reakčních podmínkách (proto také znamení). Směr chemické reakce závisí na koncentraci řídící látky na obou stranách rovnice. Pro jednoduchost se soustředíme na jednu řídící látku - totiž na vodu, která se během reakce vytváří, tj. uvolňuje. Odstraní-li se na pravé straně rovnice molekuly vody, vytvořené při kondenzaci, a tímto zásahem se zmenší jejich koncentrace v reakční směsi, probíhá reakce doprava a udrží se teoretický výsledek peptidů, který byl dosažen reakcí. Je možno ovšem postupovat i opačným směrem: přidává se reakční směsi velké množství vody (místo aby se odnímala) a tu se nevytvoří žádné peptidy, nebo jen velmi málo (podle množství vody). Místo toho obdržíme výchozí látky reakce, tj. aminokyseliny (srov. II). Je-li tedy v reakční směsi přebytek vody, nedochází k syntéze peptidů, nastane rovnováha výchozích látek-aminokyselin, jež jsou stavebním materiálem živé buňky. Tento jev se nazývá zákon o působeni mas a je platný při všech reverzibilních reakcích (vratných, jež probíhají i zpětně). Krátce řečeno: při reakcích uvedeného druhu nedojde k syntéze polypeptidů z aminokyselin, je-li k dispozici nadbytek vody.
Důsledek této všeobecně známé skutečnosti organické chemie je důležitý: Koncentrované aminokyseliny se v nějakém praoceánu, kde je nadbytek vody, vůbec nespojí v peptidy, anebo jen stěží. A jestli se nějaké polypeptidy vytvoří, tak je nadbytečná vodní masa rozloží na jejich výchozí látky (aminokyseliny). Proto je praoceán tím nejposlednějším místem na zeměkouli, kde by mohly samovolné vzniknout z aminokyseliny bílkoviny-nositelé života. Přesto skoro všechny učebnice přírodopisu uvádějí tento hrubý omyl, aby nějak zdůvodnily vývojovou teorii a biogenezi. Takový člověk musí znát organickou chemii opravdu špatně, nebo ji vědomě ignorovat, když nebere v úvahu uvedené skutečnosti. Nebo je tak ohlušený materialistickou neodarwinistickou filosofií, že zapomíná na základní fakta přírodních věd a chemie, či je přehlíží. K tomu se ale ještě vrátíme.
3. Vlastnosti stavebního materiálu života:
aminokyseliny a jejich chiralita
Je to mimo jiné 20 aminokyselin, co tvoří z čistě hmotného hlediska, základní stavební materiál živé buňky. Bez nich by nemohl život, jak jej dnes známe, ani vzniknout, ani existovat. Aminokyseliny sice mohou za jistých podmínek v praatmosféře vzniknout za pomoci blesku, avšak rozhodně a zásadně není pravda, i když to dnes tvrdí mnozí neodarwinisté, že ony náhodou vzniklé aminokyseliny se dají použít pro vytvoření živé protoplasmy. To je tedy vyloučeno. Millerovy aminokyseliny by byly pro samotný vznik života zcela a absolutně neschopné. Totéž platí pro všechny náhodou vzniklé substance a aminokyseliny, jež vytvářejí racemáty. Toto tvrzení je kategorické, má absolutní platnost a nelze je oslabit poukazem na mimořádné podmínky praoceánu či praatmosféry.
Nyní musíme vysvětlit proč je předešlé tvrzení oprávněné. Atom uhlíku, který je ve všech aminokyselinách, má strukturu podobnou pravidelnému čtyřstěnu, tj. geometrického tělesa ohraničeného čtyřmi rovnoramennými trojúhelníky.
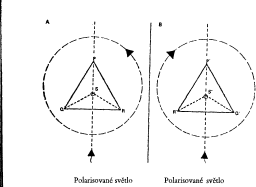
Obrázek 1. Znázornění chirality na strukturním modelu dvou molekul.
A: čtyřstěn, svislý pohled shora, znázorněn dvojrozměrně. P, Q, R, S označují hroty čtyřstěnu a znamenají čtyři mocenství atomu uhlíku. Střed základního trojúhelníku (prostorově: hrot ležící proti základnímu trojúhelníku) tvoří atom uhlíku S. Rovina polarizovaného světla se v této molekule (A) otáčí doleva (proti směru hodinových ručiček).
B: čtyřstěn, svislý pohled shora, znázorněn dvojrozměrně. Vysvětlivky jako u A; je však třeba si všimnout, že Q a R leží obráceně ve srovnání s A. Rovina polarizovaného světla se v této molekule (B) otáčí doprava (ve směru hodinových ručiček).
Čtyři mocenství atomu uhlíku jsou geometricky tak rozloženy jako čtyři hroty čtyřstěnu (P, Q, R, S). Schematicky lze tento stav znázornit následovně: (A a B), přitom P, Q, R, S znázorňují různé složky mocenství (obr. 1).
A a B mají z hmotného hlediska totožnou strukturu - jen jejich prostorové uspořádání je rozdílné. A je zrcadlovým obrazem B - a přirozeně naopak. Znázorňuje-li nyní A a B strukturní model skutečné molekuly, bude se rovina polarizovaného světla, jež je vedeno skrz A, otáčet opačným směrem, než totéž světlo, jež je vedeno skrz B.
Zrcadlo uprostřed ukazuje zrcadlové uspořádání obou molekul. Čtyřstěn A je zrcadlovým obrazem čtyřstěnu B. A proto nikdy nemůže být položen na obraz B tak, aby se P,Q,R,S jak na A tak na B úplně kryly; právě tak nelze položit levou rukavici na pravou (obě dlaní nebo obě hřbetem vzhůru), aby na sobě ležely všechny prsty a oba palce. Levá a pravá rukavice jsou zrcadlové obrazy, protože moje levá ruka je zrcadlovým obrazem pravé. Jediný rozdíl mezi A a B spočívá v prostorovém uspořádání jejich částí a ne v jejich chemickém složení, které je v obou případech - u obou zrcadlových obrazů -identické.
Tento jev zrcadlové podobnosti dvou chemicky jinak totožných molekul se označuje odborným chemickým výrazem: Chiralita. Dvě molekuly, jež jsou jedna druhé zrcadlovým obrazem, se tedy rozlišují výlučně svým prostorovým uspořádáním a ne svou chemickou analýzou. Vytvářejí-li se nyní dlouhé řetězce molekul, jež jsou různě prostorově uspořádány, tu je každému jasné, že bude jinak vypadat řetězec levotočivých molekul a jinak pravotočivých anebo ze směsi obou.. Chemické složení obou řetězců by sice bylo identické, avšak prostorové uspořádání řetězců rozdílné.
Vytvářejí-li se nyní z aminokyselin dlouhé řetězce bílkovin - jako např. bílkoviny v lidském mozku - tu hraje velkou roli prostorové uspořádání příslušných aminokyselin (jejich chiralita). Prostorové vlastnosti výchozích aminokyselin určují vlastnosti výsledných bílkovin. Pravotočivé aminokyseliny tvoří zcela rozdílné řetězce bílkovin než levotočivé nebo racemáty (opticky inaktivní směsi stejných dílů levotočivé a pravotočivé formy téže organické sloučeniny).
Ukázalo se, že bílkoviny, jež vytvářejí živou protoplasmu, jsou vesměs z levotočivých aminokyselin. Pravotočivé aminokyseliny i bílkoviny jsou v živé protoplasmě velmi řídké, neboť vytvářejí bílkoviny, jež se nehodí do látkové výměny živé buňky a často jsou jedovaté. Levá chiralita aminokyselin je tedy pro život absolutně nutná. Pravotočivé formy nemohou být v žádném případě nositeli života. Tato chiralitní zvláštnost jde však dál. Zatímco všechny aminokyseliny bílkovin života jsou levotočivé, ukazují nukleové kyseliny pravotočivost (2).
Odkud se ví, která molekula je levotočivá a která je pravotočivá? Vede-li se polarizované světlo (světelné vlny, jež kmitají v jedné rovině) roztokem levotočivých aminokyselin, otáčí se rovina polarizovaného světla doleva. Opačná chiralita stáčí rovinu polarizovaného světla doprava. Absolutní směr polarizovaného světla však nezávisí pouze na sestavení jednotlivých složek, nýbrž také na jejich chemické podstatě.
(2) Srov. např. M. EIGEN a RUTHILD WINKLER: Das Spiel, Nakl. Piper (München/Zürich 1975). Str. 142.
Z předešlého vyplývá, že aminokyseliny, jež mají sloužit biogenezi jako stavební materiál života, musí vykazovat správnou chiralitu. Aby byla možná biogeneze, musel by být všechen stavební materiál (aminokyseliny) pro živou protoplasmu levotočivý. Je velmi důležité zdůraznit tento bod - totiž že všechen stavební materiál molekuly, který se podílí na syntéze bílkovin života, musí být » opticky čistý « a » levý « - t. zn. levotočivý, bez jakékoliv příměsi pravotočivých forem. Živá protoplasma je v této věci vrcholně choulostivá: vyžaduje naprosto » čistou stravu «. I když jsou k dispozici jen malá množství pravotočivého druhu molekul aminokyseliny, vznikají bílkoviny, které mají prostorové rozdíly, a proto nejsou schopny života. Často jsou dokonce takové bílkoviny pro život zhoubné.
Dáme-li živé buňce jako potravu racemát, tj. směs 50% levotočivých a 50% pravotočivých aminokyselin, vezme si levotočivý druh a přebuduje jej na svou protoplasmu. Určité buňky mají schopnost přijmout pravotočivé formy; avšak nejprve je rozloží, přestaví na levotočivé a pak je vestaví do živé buněčné bílkoviny. Avšak buňky nemohou použít pravotočivé formy jako takové. Pravotočivé formy se z racemátu nikdy nepřetvoří na levotočivé samovolně, aniž by je živá buňka nerozložila a nepřestavěla.
Existují však zajímavé jevy, jež souvisejí s chiralitou. Jisté organizmy jako penicillium notatum (buňka, jež syntetizuje penicilín) se chrání proti vetřelcům tím, že zbuduje pravotočivé kyseliny, které usmrcují » nepřátele « otrávením. Penicilín sám je takovým biologickým jedem proti vetřelcům. Jednou jeho složkou je pravotočivá kyselina. Živá buňka si tedy » umí « poradit, jak s levotočivými, tak s pravotočivými kyselinami. Dokáže dokonce rozložit pravotočivé formy a přestavět je pro svou potřebu na levotočivé. Avšak v prabuňce se nemohly utvořit biologické bílkoviny z pravotočivých aminokyselin nebo ze směsi 50% pravotočivých a 50% levotočivých aminokyselin (tedy z racemátu). Při biogenezi nemohly vzniknout z racemátů nebo z pravotočivých kyselin bílkoviny života.
Nyní jsme s to doložit naše původní tvrzení, že prabuňka nikdy nevznikla ze směsi aminokyselin, jak tvrdí americký biochemik Stanley Lloyd Miller (ani přes stadium koacervátů, jak už před ním tvrdil Alexander Ivanovič Oparin), které se čirou náhodou utvořily v praatmosféře za pomoci blesku. Hlavní důvod tkví v tom, že všechen stavební materiál živé buňky - aminokyseliny nebo i jiné asymetrické substance (tj. schopné zrcadlového uspořádání) které jsou potřebné pro život a které vznikají pomocí blesku, tj. náhodou nebo i jinými nebiologickými procesy, tvoří výhradně racemát, tedy 50% levotočivých a 50% pravotočivých forem. Aminokyseliny, jichž dosáhl Miller jsou výlučně racemáty, a proto jsou zásadně naprosto » neschopné života «. Racemát není za naprosto žádných okolností schopen vytvořit živou bílkovinu nebo životaschopnou protoplasmu. Prostorová struktura racemátu není podle současných poznatků biochemie vůbec vhodná pro syntézu života, aby bylo možno při vzniku praživota vytvořit životaschopnou protoplasmu a vitální bílkoviny, je nutno vyjít z opticky čistých levotočivých aminokyselin. Jako pramen nestačí směsi levotočivých a pravotočivých forem. Blesk a náhoda nedokážou v žádném případě vytvořit absolutně čisté levotočivé aminokyseliny - vytvářejí výlučně racemáty, tj. 50% levotočivých a 50% pravotočivých forem.
Všechen stavební materiál při vzniku praživota musel mít čistě levou chiralitu. Avšak čirá náhoda a všechny přírodní, neživé nebo neasymetrické chemické procesy dodávají výlučně racemáty. Zde nemluvíme o enzymech, jež jsou asymetrické a opticky aktivní. Tak Miller a jeho kolegové nevyrobili v simulované praatmosféře za pomoci blesku a čiré náhody žádný vhodný stavební materiál živé buňky, nýbrž výhradně života neschopné racemáty. Současná přírodní věda nezná žádnou metodu, která by racemáty náhodným anorganickým procesem rozdělila na levotočivé a pravotočivé formy. Blesk, praatmosféra a náhodné anorganické procesy nemohou dokázat žádné takové optické rozdělení, a to jak z teoretického, tak ani z experimentálního hlediska. Mezi jednoduchými levotočivými a pravotočivými aminokyselinami neexistují žádné chemické rozdíly. Existují jedině prostorové rozdíly. Tady čirá náhoda nemůže zásadně pomoci při optickém dělení, jež je nutné pro syntézu života z aminokyselin. Aby se dalo opticky dělit, musí se zavést do reakčního systému nějaké už existující opticky aktivní centrum jako bruzin, strychnin či podobné substance. Avšak čirá náhoda nikdy nevytváří nějaké opticky aktivní centrum - už proto nemůže náhoda nikdy dát vznik živé buňce.
Z tohoto důvodu je přírodovědeckým omylem tvrzení, že Millerovy experimenty prokázaly možnost života za pomoci čistě přírodních pochodů (organická chemie) a čiré náhody. Je to vědomě klamající polopravda, protože Miller a jeho kolegové po něm vyrobili jen racemáty, které jsou pro biogenezi tak nepotřebné jako žádná jiná aminokyselina. Je totiž nezměrně těžší a vyžaduje to mnoho umu a vědomostí syntetizovat čisté formy nějaké aminokyseliny než její racemát.
Už více než 80 let se věhlasní přírodovědci marně snaží nějakým způsobem za pomoci čistě anorganického procesu a bez použití živých buněk vyrobit opticky čisté aminokyseliny náhodným anorganickým postupem. Kdyby někdo dokázal ulomit ostří tomuto problému, jenž je základním pro vyřešení záhady biogeneze, stal by se vážným kandidátem na udělení Nobelovy ceny. Vždyť právě tento bod je hlavní překážkou, na níž ztroskotává věrohodnost běžných materialistických teorií o samovolném vzniku života. Z tohoto důvodu se také v dnešních učebnicích zamlčuje tato velká obtíž chirality. Jejich autoři jsou totiž často zastánci materialistického názoru na biogenezi. Vědí příliš dobře, že problém racemátů nezměrně ztěžuje samovolný vznik života za pomoci čiré náhody a nahodilých přírodních chemických procesů. Proto význam tohoto jevu zamlčují nebo jej povrchně odbývají.
Různí přírodovědci se po léta pokoušeli opticky dělit racemáty (odlučovat levotočivé od pravotočivých forem) tím, že vytvářeli aminokyseliny na opticky aktivním povrchu za pomoci nahodilého chemického procesu. Jako opticky aktivní povrch byly zvoleny quarzové krystaly, které jsou opticky aktivní bez součinnosti atomového asymetrického centra, avšak marně. Zkoušelo se vytvořit opticky aktivní aminokyseliny pod vlivem kruhového polarizovaného světla. Byla tu domněnka, že by se kruhovým polarizovaným světlem jedna optická forma rychleji zničila než druhá, a tak by se dospělo k oddělení izomerů. A přesto proběhly všechny pokusy více méně marně, byla zjištěna jen mizivě malá optická aktivita. Musíme si uvědomit, že pro samovolnou syntézu živé buňky by bylo zapotřebí více méně stoprocentní optické čistoty u asi 20 různých aminokyselin. Za pomoci pouze anorganické nahodilé reakce nebylo nikdy možno dosáhnout opticky čisté formy. Z tohoto důvodu a ještě z jiných je samovolný vznik života dodnes experimentálně nemožný. Teoretické úvahy jen dotvrzují tento negativní nález na základě pokusů.
Můžeme tedy jen konstatovat, že nelze syntetizovat vlastní stavební materiál živé buňky, tj. asi 20 opticky čistých aminokyselin, anorganickými nahodilými procesy. Millerovy pokusy mají proto velmi málo společného se skutečným vznikem života, i když je učebnice líčí jako poslední článek řetězce důkazů, že život na zemi vznikl čirou náhodou. Tvrdit, že Miller dokázal první krok k biogenezi, je vědomé klamání nezasvěcené veřejnosti. Vlastní problém se zamlčuje, aby se materialistická filosofie stala přírodovědecky přijatelná. Je to hrubá manipulace přírodních věd v zájmu běžných materialistických filosofií.
Opticky čisté levotočivé aminokyseliny je ovšem možné vyrobit v laboratoři. » Vyrobil jsem celé kilogramy opticky čistého alaninu, kyseliny alfa-aminomáselné a mnoho jiných sloučenin, ale nikdy čirou náhodou. Za pomoci vědomostí (know-how) se to daří, nikdy však čirou náhodou (opak know-how). Je k tomu zapotřebí složitých chemických pochodů, aby se dosáhl tak úctyhodný výsledek. Pro takové oddělení je zapotřebí mít už existující aktivní centrum, a to nevznikne nikdy náhodou. Každý vzdělaný chemik se jen usmívá, když někdo mluví o čiré náhodě v této oblasti. «
4. Jak vysvětluje Manfred Eigen, odkud pochází chiralita potřebná pro biogenezi
Z nějaké racemátové směsi kyselin nemůže vzejít žádná přímá samovolná biogeneze, protože nemá k dispozici chiralitu životně nutných bílkovin. Prostorově podmíněný systém buněčných receptorů by nefungoval. Kdyby však přesto vznikla živá buňka z racemátové směsi, měla by pramálo společného s životními formami, jaké známe dnes. Vždyť život, jak jej známe dnes, závisí na chiralitě a na prostorově podmíněných receptorech, které jsou opět ve své funkci závislé na chiralitě.
Eigen se ptá, proč nedochází v přírodě k pokojné koexistenci levotočivých a pravotočivých forem života. (3) Proč se objevují výlučně levotočivé formy v aminokyselinách a výhradně pravotočivé v nukleových kyselinách? Pro každého biologa je odpověď jasná: Život je absolutně závislý na prostorových strukturách a na chiralitě. Eigenovy odpovědi na tento problém jsou zajímavé: a) Je to charakteristickým rysem systému, který se sám reprodukuje, píše, že obě optické formy nemohou koexistovat. Ačkoliv je tato věta naprosto přesná, laikovi přirozeně mnoho nevysvětlí. b) Protože přírodní výběr rozhoduje o přežití na základě » všechno nebo nic «, píše Eigen, je prý to jen otázka času, která forma se prosadí, zda levá či pravá.
Tato výpověď je důležitá, neboť tím Eigen řekl něco, s čím může souhlasit jen málo molekulárních biologů.
(3) Eigen a Winkler, uvedené dílo (pozn. 2), strana 114 a násl.
Chce tím tvrdit, že levotočivé a pravotočivé aminokyseliny a jejich směsi byly dříve životaschopné. To je ovšem filosofické tvrzení, avšak žádná přírodní věda, neboť experimentálně dnes nežije žádná racemátová buňka. To také potvrzuje Eigen pod a). Podle svého vyjádření v b) by se měly podílet na samovolné biogenezi racemáty a pravotočivé bílkoviny. Kdyby tomu tak bylo skutečně, reagoval by » racemátový život « nebo » pravý život « sám na sebe i vůči prostředí docela jinak než » levý život «, neboť vztahy droga - receptor (speciální orgán citlivý na určité podněty, změny prostředí) nebo agonista - antagonista by byly různorodou chiralitou tak narušeny, že by musela být přebudována celá buněčná látková výměna. Nemohlo by se tedy jednat o tytéž druhy života, o nichž jednáme dnes. Bod a) je z každého hlediska správný - je to charakteristickým rysem života, že obě formy nemohou pokojně koexistovat. Jakpak by však mohl vzniknout život na nějakém místě z racemátové směsi? Veškeré prostorové vztahy v oblasti agonista antagonista by se staly chaotické pod vlivem nejednotné chirality. Bod b) předpokládá, že »racemátový život« pomocí samovolné biogeneze produkuje stejné druhy a rody, jako jsou dnešní. To by bylo opravdu problematické. Vždyť 1. racemátový život experimentálně neexistuje. 2. Stěží může existovat teoreticky. 3. Přírodní výběr. který má od sebe oddělit levotočivé formy od pravotočivých, může nastat teprve tehdy, až existuje racemátový život.
Z Eigenových výpovědí vyvozujeme dva závěry. Za prvé: je naprosto správné, jak tvrdí Eigen, že charakter systému, který je schopen sám sebe reprodukovat, nepřipouští žádné setření hranic chirality. Buněčný systém látkové výměny, který plně funguje a sám sebe reprodukuje, by tedy jistě nemohl trpět žádnou pokojnou koexistenci pravotočivých a levotočivých forem. Racemáty by nemohly vyvolat žádnou biogenezi, jakou známe, neboť prostorové (stereochemické) podmínky, na nichž závisí život, který známe, by nebyly splněny. Druhý závěr zní: protože neexistuje žádný život, jak jej známe na základě racemátů, nemůže být řeč o nějakém přírodním výběru v životě, který by časem rozhodl, zda v životním boji zvítězí pravá nebo levá forma. Kde neexistuje život, tam samozřejmě neexistuje přírodní výběr. Jestliže racemáty jako takové nemohou být nositeli života, nemůže žádný přírodovědec tvrdit, že přírodní výběr způsobil optické dělení v tomto »neživotě«. Když při nějakém závodu všichni běžci leží mrtvi na startovní čáře, jak lze mluvit vůbec nějakém vítězství, dosaženém výběrem závodní soutěže. Proto výběr, který se zakládá na racemátech, nemůže vykouzlit žádné optické dělení, a proto k němu nikdy nedošlo. Vysvětlení o původu optické aktivity života jsou proto tautologické.
Ve světě pohádek vznikají všelijaké zamotané situace. Čarodějnice zabije prince nebo krásná, milovaná princezna umře na záhadnou nemoc, kterou na ni uvalila zlá čarodějnice. Všechny tyto problémy vždy vyřeší kouzelná hůl. Někdo ji přinese, dotkne se jí zraněného místa nebo mrtvého těla a hle, všechno je zase v pořádku! Mrtvá princezna stojí před námi v oslňující kráse a ze zlé čarodějnice zbude jen kouř a sirný zápach.
Podobně i současná biologie objevila kouzelný proutek, který vyřeší jediným mávnutím všechny biologické i chemické procesy. Je třeba vysvětlit, jak vzniklo závratně komplikované ústrojí proteinové molekuly?
Je třeba vysvětlit, jak vznikají optické izoméry? Chce někdo vědět, proč jistý druh motýlů má na svých křídlech obraz orlího oka? Kouzelný proutek, který se nazývá přírodní výběr, vysvětlí všechny tyto zázraky beze zbytku. Vysvětlí, jak vznikla nezměrně složitá biologická ústrojí - proteinových molekul. Je to ale opravdu pohádkové vysvětlení - stroje vznikají samy od sebe, samočinně, jako mávnutím kouzelného proutku. Vysvětluje, jak se vytvořily biliony elektrických kontaktů v mozku. Vysvětluje nepředstavitelně složité propojení komputeru, který nazýváme mozek. Tento přelud vyvěrá z jediného chybného poznatku: Domnívá se, že konkurenční boj automobilových továren o nové trhy vyrábí nové typy aut. Velkoryse se přitom zapomíná, že konkurenční boj pouze třídí plány a koncepty, ale nevytváří nové návrhy a nové výrobky. Přirozený výběr má provést optické dělení i tam, kde není žádný život a žádný výběr. Přírodní výběr je takovým moderním kouzelným proutkem!
Stojíme před jedním z největších nevyřešených problémů biologie, které dnes ještě existují. Bílkoviny života musí mít ke svému vzniku opticky čisté levotočivé aminokyseliny. Musí z nich vycházet biogeneze, zvláště má-li být spontánní, aby dodala bílkoviny života, jaké známe. Podobným způsobem se musely vytvořit i pravotočivé nukleové kyseliny. Přirozený výběr - navzdory dnes běžným tvrzením - nemůže hrát žádnou roli při biogeneticky nutném oddělení pravotočivých a levotočivých forem. Vždyť před vznikem života nemohl existovat žádný přírodní výběr!
Proto se musíme vrátit k původní otázce: Odkud » vzal « proces vzniku života opticky čisté aminokyseliny, aby vytvořil první opticky čisté bílkoviny a nukleové kyseliny? Je naprosto jisté, že aminokyseliny, které syntetizoval Miller a jeho kolegové za pomoci blesku a čiré náhody v praatmosféře, by se pro biogenezi vůbec nedaly použít - ještě méně než automobilové součástky na výrobu velmi přesných hodinek -, neboť to všechno byly racemáty.
Laboratorní pokus nám ovšem může dát poznat, jak se dojde k opticky aktivním aminokyselinám, jež jsou nutné pro biogenezi. Experiment nám má v našich představách dát vysvětlení, jakým způsobem vznikl život. Proto se nyní zaměříme na experiment, který nám poskytuje jedinou pevnou půdu pro teoretické úvahy. Chceme-li v laboratoři vyrobit z racemátů optickou aktivitu, musíme především do racemátového systému vestavět opticky aktivní centrum, opticky aktivní molekulu. Racemátová kyselina se sloučí s levotočivou nebo pravotočivou bází jako je strychnin nebo brucin. Tímto způsobem se vytvoří dvě rozdílné látky, které se dají rozdělit: a) levotočivá kyselina/levotočivý brucin; b) pravotočivá kyselina/levotočivý brucin. Látky a) a b) mají často rozličnou rozpustnost, takže je lze oddělit pomocí krystalizování. Když jsme a) a b) rozdělili krystalizováním, rozložíme je opět na jejich jednotlivé složky: a) dodá levotočivou kyselinu a brucin; b) dodá pravotočivou kyselinu vedle brucinu.
Aby se dal racemát rozložit na jeho levotočivé a pravotočivé složky, je nutno zavést do racemátového systému 1. molekulu, která vykazuje už předtím existující optickou aktivitu; 2. použít know-how, aby se uskutečnilo dělení, a 3. dodat energii.
Živá buňka dokáže vyrobit velká množství opticky aktivních látek. Má totiž asymetrická centra, která existovala již před tímto procesem. Zvířata i rostliny mají takovou schopnost. Vlastní totiž na svém genetickém kódu informaci nebo » vědět jak « zbudovat opticky aktivní látky. Buněčná látková výměna dodává energii. Tak se experimentálně vědecky dosáhne optická aktivita. Know-how ve formě kódu nebo enzymu je vedle energie nezbytné pro tento postup optického dělení. A k tomu ještě musí být po ruce předem existující optická aktivita. Tuto předem existující optickou aktivitu dodávají enzymy a genetický kód dodává » vědět jak «. Avšak ani optická aktivita, ani know-how nevznikly čirou náhodou.
To všechno ale neřeší náš problém - odkud pocházely opticky čisté aminokyseliny pro syntézu první buňky při biogenezi. Eigenův návrh, že biologie začala s racemáty - » s racemátovým životem « - většina biologů nepřijímá, jak jsme už konstatovali. Proto musela optická aktivita vzniknout před biogenezí, a ne po ní. Dejme tomu, že praživot vzešel z aktivních látek, které bylo možno dodat, než se objevil život. Výroba optické aktivity vyžaduje energii, již předem existující aktivní centrum a » vědět jak «. Před biogenezí nejsou také žádným problémem zdroje energie. Odkud však vezmeme v anorganickém světě optické aktivní centrum a know-how před biogenezí?
Problém se dá formulovat ještě jednodušeji: Pro experimentální výrobu optické aktivity, jak víme, jsou nezbytně nutné tři složky: know-how, energie a již existující aktivní centrum. Toto centrum můžeme ovšem za určitých experimentálních podmínek postrádat -ačkoliv se to v běžné laboratorní praxi nedělá. Když Pasteur pracoval s kyselinou vinnou, zjistil, že některé její krystaly mají určitou formu a jiné opět odlišnou. Oddělil tedy od sebe obě formy tím, že je dal pod mikroskop a odděloval je ručně pomocí pinsety. Použil tedy své oko - a svou inteligenci - aby je rozdělil na základě rozdílných tvarů - je to příklad rozlišování vzorků (pattern recognition) na základě know-how. Rozdílná podoba krystalů vinné kyseliny pocházela ze dvou asymetrických center v racemátové molekule vinné kyseliny. V případech, kde je pouze jediné centrum v molekule, by nebyla možná Pasteurova dělicí metoda.
Když se tedy provedlo optické dělení na základě rozpoznávání rozličných podob, mohla by se takto získaná optická aktivita použít k tomu, aby se dělily jiné racemáty podle běžné laboratorní praxe.
Právě jsme vytvořili předpoklad (předem existující optickou aktivitu) pro optické dělení - a to za pomoci » vědět jak «. Předem existující optická aktivita se ukazuje jako absolutně nutná - inteligenci nebo know-how je možno v určitých případech nahradit.
Nyní jsme s to pustit se do řešení našeho prapůvodního problému. Odkud pocházejí opticky aktivní stavební bloky (opticky aktivní aminokyseliny) biogeneze? Know-how a energie by je mohly dodat. Energie není žádným problémem. Avšak zařadit do biogeneze vedle hmoty a energie jako třetí složku nezbytné » vědět jak «, dělá velké těžkosti člověku, který je ve vleku materialistické filosofie. Hmota, čas a prostor v anorganickém světě (sféra materialismu) nemají v sobě žádné know-how, žádnou inteligenci, žádnou inteligenci. Abychom však dokázali oddělit různé formy, abychom dokázali vybudovat složité stroje, jakými jsou molekuly bílkovin, musíme vědět, jak na to, musí tu být inteligence, to znamená, že musíme mít plán, inteligenci, koncept, chceme-li dosáhnout optické aktivity. Bez » vědět jak «, bez inteligence to prostě nejde.
Protože v našich dimenzích času a prostoru není místa pro know-how (či inteligenci), které jsou naopak nezbytné pro vznik života a pro vytvoření optické aktivity stejně jako je absolutně a bezpodmínečně nutné pro výrobu všech strojů včetně molekul bílkovin a enzymů, mám za to, že » vědět jak «, koncept, inteligence hrály určitou roli při prapůvodním optickém dělení. O zdroji tohoto nezbytného know-how je možno mít různé názory. Materialista Aldous Huxley si představoval nějaký universální » Think-tank «, aby se nějak dostal přes toto úskalí. K tomu se ještě později vrátíme.
» Vědět jak « a inteligence jsou přirozeně úzce spjaty. Je jich zapotřebí nejen k optickému rozdělení izomérů, nýbrž stejně i k výrobě strojů všeho druhu. Vždyť stroj podle definice je inteligentní zařízení. Manfred Eigen sám pěkně ukazuje, že živé bílkoviny jsou právě tak stroje, jako automobil je stroj - inteligentní stroj : Proteiny se právem považují za nejmenší nám známé stroje. Řežou, svářejí, vyměňují, třídí, převážejí, předělávají a přitom každá proteinová molekula, po případě enzym, slouží zcela vyhraněnému účelu. Jacques Monod je proto nazývá: teleonomické struktury. Jejich výstavba se neřídí nějakými estetickými měřítky. Jediným požadavkem je účelnost, stejně jako u nějakého stroje, který vymyslel, navrhl a vyrobil člověk. (4)
(4) Cit. EIGEN a WINKLER: Das Spiel (pozn. 3), Str. 146.
1. Bílkovinné molekuly (proteiny, enzymy) jsou tedy především inteligentní struktury, složité stroje, právě takové, jako by je vymyslel člověk. Ono » být vymyšlen « vnáší do biogeneze, do představy o vzniku života inteligentní účelnost, koncept, » vědět jak «. Optické dělení vyžaduje chemické know-how, stejně jako konstrukce kteréhokoliv stroje. I bílkovinné stroje vyžadují projekt, inteligenci.
2. Z těchto důvodů nemůže dnes žádný opravdový a čestný přírodovědec věřit na pravznik života čirou náhodou, tj. bez inteligence. Základem biogeneze není čirá náhoda, nýbrž právě její protipól: koncept, projekt či » vědět jak «. Stavba bílkovinnových strojů, stejně jako rozdělení optických protinožců (tj. levotočivých a pravotočivých forem) vyžadují pravý opak náhody, který nazýváme: vědět jak. Kdo myslí v kategoriích experimentů a zákonů přírodních věd, je nucen udělat tento závěr. Čistě přírodovědecké metody však ještě nedokázaly vysvětlit, odkud pochází to nezbytné » vědět jak « či tato inteligence. Známe však hmotu a dimenze času a prostoru tak dobře, že je nepravděpodobné, aby se potřebné » vědět jak « skrývalo v čistě hmotném časoprostorovém kontinuu. Druhý termodynamický zákon, jenž říká, že hmota ponechána sama sobě se neorganizuje, vylučuje, že by ve hmotě byl nějaký sklon k uspořádání či projekt vyšší organizace, tedy inteligence. Proto jsem toho názoru, že koncept, » vědět jak « a inteligence se nenajde pouze v lidském vědomí, nýbrž i mimo naše časoprostorové kontinuum.
Budeme stěží s to přijít na mechanizmus biogeneze a biologie, pokud budeme připisovat stavbu biologických bílkovinných strojů a optické dělení aminokyselin čiré náhodě, místo jejímu protipólu (know-how). Totéž dilema se vynořuje v dnešní biologii tam, kde se snaží připisovat vznik genetického kódu jen čiré náhodě. Toto dilema a jeho následky jsem popsal v jiné své knize. (5)
(5) A. E. WILDER SMITH: Demission des wissenschaftlichen Materialismus, Nakl. Telos Neuhausen-Stuttgart 1976.