Původ rostlin
(The
Origin of Plants)
Harun
Yahya
http://www.signsofcreation.com/refuted_12.htm
Přeložil
M. T. – únor 2008
Vědci dělí život na zemi do pěti (nebo
někdy šesti) říší. Dosud jsme se soustředili hlavně na
největší říši, říši zvířat. /viz webové stránky/
V předešlých kapitolách jsme se zamýšleli nad původem
života jako takového, studovali jsme proteiny (bílkoviny),
genetickou informaci, strukturu buňky a bakterie, tedy problémy
týkající se dvou dalších říší, Prokaryotae a Protista.
Dnes však existuje jiné důležité téma, na něž se musíme
soustředit – původ říše rostlin – Plantae.
V původu rostlin nalézáme stejný obraz
jako byl ten, s nímž jsme se setkali, když jsme zkoumali
původ zvířat. Rostliny mají velmi komplexní struktury, a
není možné, aby tyto struktury vznikly náhodnými událostmi
a aby se rostliny vyvíjely jedna v druhou. Fosilní nálezy
ukazují, že různé třídy rostlin se vynořily do světa
vždy náhle, každá se svou specifitou, a bez známek evoluce.
Původ rostlinné buňky
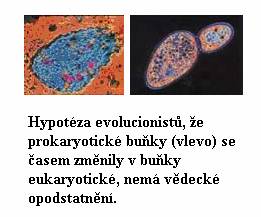
Podobně jako buňky živočišné, i buňky
rostlinné patří k typu známému jako „eukaryotní“.
Nejpodstatnějším rozlišujícím rysem těchto buněk je to,
že mají buněčné jádro, a molekula DNA (v níž je
zakódována jejich genetická informace) leží v tomto
jádře. Na druhé straně, někteří jednobuněční tvorové
jako třeba bakterie buněčné jádro nemají, a molekula DNA je
umístěna volně v buňce. Tento druhý typ buňky nazýváme
„prokaryotní“. Tento typ buněčné struktury s volnou DNA
neuzavřenou do jádra je ideálním pro bakterie, protože
umožňuje velmi důležitý proces – z hlediska bakterie –
transferu plazmidů (to znamená transferu DNA z buňky do
buňky).
Plazmid = kruhová molekula DNA nesoucí
jeden nebo několik genů, která může existovat v bakteriích
mimo vlastní chromozom a nezávisle na něm se množit. Plazmid
může být mezi bakteriemi předáván (viz konjugace). Někdy
obsahuje geny rezistence proti některým antibiotikům. (pozn.
překl.) .
Protože evoluční teorie musí sestavovat
živé organizmy v řadě „od primitivního ke složitému“,
předpokládá, že prokaryotní buňky jsou primitivní, a že
se eukaryotní buňky vyvinuly právě z nich.
Než začneme probírat neplatnost tohoto
tvrzení, bude užitečné ukázat, že prokaryotní buňky
nejsou vůbec „primitivní“. Bakterie má asi 2000 genů;
každý gen obsahuje okolo 1000 písmen (spojení). To znamená,
že informace v bakteriální DNA měří asi 2 milióny
písmen. Podle téhle kalkulace se informace v DNA jedné
bakterie rovná 20 románům, každému po 100 000 slovech (326).
Každá změna v informaci v kódu DNA bakterie by byla tak
zhoubná, že by to vyřadilo veškerou činnost bakterie. Jak
jsme viděli, chyba v bakteriálním genetickém kódu znamená,
že bakterie nebude fungovat – tj., buňka zemře.
Kromě této citlivé struktury, která
nevzniká náhodnými změnami, činí tvrzení evolucionistů
neopodstatněným též fakt, že nebyla nalezena „přechodná
forma“ mezi bakterií a eukaryotními buňkami. Například
slavný turecký evolucionista profesor Ali Demirsoy přiznává
nepodloženost scénáře, že se bakteriální buňky vyvinuly
v buňky eukaryotní a pak ve složité organizmy vystavěné
z těchto buněk:
„Jednou z nejobtížnějších fází
k vysvětlení v evoluci je vědecky vysvětlit, jak se
organely a složité buňky vyvinuly z těchto primitivních
tvorů. Mezi dvěma hlavními formami života nebyla nalezena
žádná forma přechodná. Jedno- i vícebuněční tvorové
mají všichni tuto komplikovanou strukturu, a ještě nebyl
nalezen žádný tvor či skupina tvorů s organelami
jednodušší konstrukce v jakémkoli ohledu či vůbec
primitivnější. Jinými slovy, děděné organely se vyvinuly
právě takové, jaké jsou. Nemají jednoduché nebo primitivní
formy“ (327).
Člověk se diví, co přimělo profesora
Aliho Demirsoye, loajálního přívržence evoluční teorie,
k tomu, že učinil tak otevřené přiznání? Odpověď na
tuto otázku můžeme dát zcela jasně, prozkoumáme-li velké
strukturální rozdíly mezi bakteriemi a rostlinnými buňkami.
Ty zahrnují:
1 – Zatímco stěny bakteriálních
buněk tvoří polysacharid a bílkovina, stěny buněk
rostlinných tvoří celulóza, zcela odlišná struktura.
2 – Zatímco rostlinné buňky
mají mnoho organel, uzavřených v membránách a majících
velmi složité struktury, buňky bakteriální typické organely
postrádají. V bakteriálních buňkách jsou pouze volně se
pohybující nepatrné ribozomy. Ribozomy v rostlinných
buňkách jsou však větší a jsou připevněny k buněčné
membráně. Dále, syntéza proteinů probíhá v oněch dvou
typech ribozomů rozdílným způsobem.
3 – Struktury DNA v rostlinných a
bakteriálních buňkách se liší.
4 – Molekula DNA v rostlinných
buňkách je chráněna dvouvrstvou membránou, zatímco DNA
v buňkách bakteriálních je uložena volně v buňce.
5 – Molekula DNA v bakteriálních
buňkách se podobá uzavřené smyčce; jinými slovy, je
kruhová. V rostlinách je molekula DNA lineární.
6 – Molekula DNA v bakteriálních
buňkách nese informaci patřící pouze jedné buňce, ale
v buňkách rostlinných molekula DNA nese informaci o celé
rostlině. Například všechny informace o ovocném stromu –
jeho kořenech, kmeni, listech, květech i plodech – se
nalézají společně v DNA v jádře pouhé jediné buňky.
7 – Některé druhy bakterií jsou
fotosyntetické, jinými slovy, provádějí fotosyntézu. Ale na
rozdíl od rostlin není ve fotosyntetických bakteriích
(například cyanobakteriích) chloroplast obsahující chlorofyl
a fotosyntetické pigmenty. Tyto molekuly jsou spíše ponořeny
do různých membrán kolem celé buňky.
8 – Biochemie tvoření
informační RNA v prokaryotických (bakteriálních) buňkách
a v eukaryotických buňkách (včetně rostlinných a
zvířecích) se podstatně liší (328).
Informační RNA = druh ribonukleové
kyseliny, která vzniká přepisem z DNA (transkripcí) a
„sestřihem“ (splicing). Z jádra je transportována do
cytoplasmy, kde se ve spojení s ribozomy účastní syntézy
bílkovin (translace). Přenáší dědičnou informaci, která
je uložena v genu, a kóduje přesné pořadí aminokyselin
v bílkovině. (pozn. překl.).
 Informační RNA hraje životně
důležitou roli v přežívání buňky. Ale třebaže
informační RNA hraje stejnou vitální roli jak
v prokaryotických tak v eukaryotických buňkách, její
biochemické struktury se liší. J. Darnell napsal toto
v jednom článku uveřejněném v časopise Science:
Informační RNA hraje životně
důležitou roli v přežívání buňky. Ale třebaže
informační RNA hraje stejnou vitální roli jak
v prokaryotických tak v eukaryotických buňkách, její
biochemické struktury se liší. J. Darnell napsal toto
v jednom článku uveřejněném v časopise Science:
Rozdíly v biochemii tvoření informační
RNA u eukaryot ve srovnání s prokaryoty jsou tak hluboké, že
svědčí o tom, že návazná evoluce prokaryotických buněk
v buňky eukaryotické se jeví nepravděpodobnou (329).
Strukturální rozdíly mezi bakteriálními
a rostlinnými buňkami (o nichž jsme uvedli několik
příkladů výše) zavedly evolucionisty do další slepé
uličky. Ačkoli mají rostlinné a bakteriální buňky
některé rysy společné, většina jejich struktur se
podstatně liší. Pravda je, že jelikož v bakteriálních
buňkách neexistují organely obklopené membránou nebo
cytoskeletonem (vnitřní sítí bílkovinných filamentů a
mikrotubulů), pak přítomnost některých velmi složitých
organel a celková organizace buňky u rostlin činí zcela
neplatným tvrzení, že rostlinná buňka se vyvinula z buňky
bakteriální.
Biolog Ali Demirsoy to otevřeně
připouští, když říká: „Složité buňky se nikdy
nevyvinuly z buněk primitivních procesem evoluce .“
Endosymbiotická hypotéza a její
neplatnost
Nemožnost toho, aby se rostlinné buňky
vyvinuly z buněk bakteriálních, nezabránila evolučním
biologům v tom, aby produkovali spekulativní hypotézy.
Experimenty je však nepotvrzují (331). Nejpopulárnější
z nich je hypotéza „endosymbiózy“.
Tuto hypotézu navrhla Lynn Margulisová
roku 1970 ve své knize Původ eukaryotických buněk. V této
knize Margulisová tvrdila, že výsledkem primitivně
pospolitého a parazitického života bakterií bylo to, že se
jejich buňky změnily v buňky rostlinné a zvířecí. Podle
téhle teorie se rostlinné buňky objevily tehdy, když byla
fotosyntetická bakterie pohlcena jinou bakteriální buňkou.
Z fotosyntetické bakterie se stal uvnitř rodičovské buňky
chloroplast. Nakonec se vyvinuly, tak či onak, organely
s vysoce složitými strukturami jako je jádro, Golgiho
aparát, endoplazmatické retikulum, a ribozomy. Tak se zrodila
rostlinná buňka.
Jak jsme viděli, tato evolucionistická
teze není ničím jiným než dílem fantazie. Nepřekvapuje
tedy, že byla kritizována vědci, kteří prováděli velmi
důležitý výzkum na tomto poli s řadou témat: Můžeme
citovat D. Lloyda (332), M. Graye a W. Doolittlea (333), a R.
Raffa a H. Mahlera jako příklady takových vědců.
Endosymbiotická hypotéza je založena na
faktu, že mitochondrie živočišných buněk a chloroplasty
buněk rostlinných obsahují vlastní DNA, jinou než je DNA
v jádře rodičovské buňky. Takže, vzhledem k této
skutečnosti, je navrhováno, že mitochondrie i chloroplasty
byly kdysi samostatnými, volně žijícími buňkami.
Budeme-li však studovat chloroplasty
podrobně, ukáže se, že toto tvrzení je neprokázané.
Četné námitky činí endosymbiotickou
hypotézu neplatnou:
1 – Pokud by zejména chloroplasty
byly kdysi nezávislými buňkami, pak by došlo k jedinému
výsledku, pokud by byl chloroplast pohlcen větší buňkou:
totiž, byl by stráven rodičovskou buňkou a využit jako
potrava. Musí tomu tak být, protože i když připustíme, že
příslušná rodičovská buňka pozře chloroplast z vnějšku
omylem (místo aby jej úmyslně ulovila jako potravu), trávicí
enzymy v rodičovské buňce by jej nicméně zničily.
Samozřejmě, někteří evolucionisté obejdou tuto překážku
tvrzením „Trávicí enzymy vymizely“. Ale tohle je jasný
protimluv, protože pokud by trávicí enzymy buňky zmizely, pak
by buňka zahynula nedostatkem potravy.
2 – Ale přece jen, představme si,
že vše toto nemožné popsané v bodě 1 se stalo a že buňka,
o níž tvrdíme, že je předkem chloroplastu, byla pozřena
buňkou rodičovskou. V tomto případě čelíme dalšímu
problému: Programy všech organel v buňce jsou zakódovány
v DNA. Pokud by rodičovská buňka hodlala použít jiné
buňky, které spolkla, jako organely, pak by bylo třeba, aby
veškeré informace o nich byly již přítomny a zakódovány
v její DNA. DNA buněk spolknutých by musela obsahovat
informaci patřící buňce rodičovské. Nejen že je taková
situace nemožná, ony dvě složky DNA patřící rodičovské
buňce a spolknuté buňce by musely být kompatibilní i
v budoucnu, což je také jednoznačně nemožné.
3 – V buňce vládne velká
harmonie, kterou náhodné mutace nemohou vysvětlit. Buňka
obsahuje více než pouhý jeden chloroplast a jednu
mitochondrii. Jejich počet stoupá a klesá podle úrovně
aktivity buňky, právě tak jako počet jiných organel.
Existence DNA v tělech těchto organel je též užitečná
v reprodukci. Když se buňka dělí, všechny ty chloroplasty
se dělí také, a buněčné dělení probíhá v kratším
čase a pravidelněji.
4 – Chloroplasty jsou generátory
energie s absolutně životní důležitostí pro rostlinnou
buňku. Pokud by tyto organely nevyráběly energii, mnoho
buněčných funkcí by nepracovalo, což znamená, že by buňka
nemohla žít. Tyto funkce, jež jsou tak důležité pro buňku,
probíhají díky proteinům syntetizovaným v chloroplastech.
Ale DNA vlastní chloroplastům nestačí na to, aby tyto
proteiny syntetizovala. Větší část proteinů je
syntetizována díky rodičovské DNA v buněčném jádře
(334).
Vezmeme-li v úvahu, že situace
předpokládaná endosymbiotickou hypotézou se objevuje díky
procesu pokusu a omylu, jaké důsledky by měla v DNA
rodičovské buňky? Jak jsme viděli, jakákoli změna
v molekule DNA v každém případě nepřináší pro
příslušný organizmus přínos; naopak, jakákoli taková
mutace by jistě byla škodlivá. Ve své knize Kořeny
života vysvětluje Mahlon B. Hoagland tuto situaci:
Vzpomeňte si, že jsme se učili, že
téměř vždy je změna v DNA pro organizmus zhoubná; to
znamená, že vede k omezení schopnosti přežít. Analogicky
vzato, náhodné přídavky vět do Shakespearových her by
těžko znamenaly jejich vylepšení!… Zásada, že změny
v DNA jsou škodlivé tím, že omezují vyhlídky na
přežití, se vztahuje jak na případ, že změna v DNA je
způsobena mutací, tak na případ, že ji způsobí nějaké
cizí geny, které do ní úmyslně přidáme (335).
Tvrzení pronášená evolucionisty nejsou
založena na vědeckých experimentech, protože žádná taková
věc jako fakt, že by jedna bakterie spolkla druhou, nebyla
nikdy pozorována. Ve své recenzi další knihy Margulisové, Symbióza
v evoluci buňky, popisuje molekulární biolog P. Whitfield
situaci takto:
Prokaryotická endocytóza je buněčným
mechanizmem, na němž celý problém S.E.T. (seriální
endosymbiotické teorie) zřejmě končí. Kdyby jeden prokaryot
nemohl pohltit jiný, pak je těžké si představit, jak by
endosymbiózy mohly být prováděny. Bohužel pro Margulisovou a
S. E. T., neexistují žádné moderní příklady prokaryotické
endocytózy nebo endosymbiózy…(336).
Endocytóza = aktivní proces, jímž buňka
pohlcuje větší částice. K endocytóze patří zejména
fagocytóza a pinocytóza. Opak: exocytóza. (pozn. překl.).
Původ fotosyntézy
Další záležitostí ohledně původu
rostlin, jež vnáší do evoluční teorie strašlivý zmatek,
je otázka, jak rostlinné buňky začaly provádět
fotosyntézu.
Fotosyntéza je jedním ze základních
životních procesů na zemi. Díky svým chloroplastům
produkují rostlinné buňky škrob za použití vody,
kysličníku uhličitého a slunečního světla. Zvířata
nejsou schopna produkovat své vlastní živiny, a musí místo
toho využívat škrob z rostlin ke své výživě. Z tohoto
důvodu je fotosyntéza základní podmínkou složitého
života. Ještě zajímavější stránkou celé věci je fakt,
že tenhle složitý proces fotosyntézy nebyl dosud plně
pochopen. Moderní technologie ještě nebyla schopna odhalit
všechny jeho podrobnosti, natož pak ho reprodukovat.
 Je možné, aby tak složitý proces jako
je fotosyntéza vznikl přirozenou cestou, jak tvrdí evoluční
teorie?
Je možné, aby tak složitý proces jako
je fotosyntéza vznikl přirozenou cestou, jak tvrdí evoluční
teorie?
Podle evolučního scénáře rostlinné
buňky za tím účelem, aby mohly provádět fotosyntézu,
spolkly buňky bakteriální, které to už uměly, a přeměnily
je v chloroplasty. Takže, jak se bakterie naučila provádět
tak komplikovaný proces jako je fotosyntéza? A proč nezačala
provádět takový proces předtím? Jako je tomu i u jiných
otázek, tento scénář nemá vědeckou odpověď. Podívejme se
na to, jak evolucionistická publikace na tuto otázku
odpovídá:
Hypotéza heterotrofie navrhuje, že
nejranější organizmy byly heterotrofní a živily se polévkou
z organických molekul v primitivním oceánu. Když tihle
první heterotrofové zkonzumovali přítomné aminokyseliny,
bílkoviny, tuky i cukry, živinová polévka se vyčerpala a
nemohla už uživit rostoucí populaci heterotrofů…Organizmy,
které by mohly využívat alternativní zdroj energie, by byly
měly velkou výhodu. Pomysleme na to, že Země byla (a stále
je) zaplavována sluneční energií, kterou vlastně tvoří
různé formy záření. Ultrafialové záření je zničující,
ale viditelné světlo je bohaté na energii a neškodné.
Takže, když se organické složky
stávaly stále vzácnějšími, již přítomná schopnost
využívat viditelné světlo jako alternativní zdroj energie
mohla umožnit takovým organizmům a jejich potomkům
přežívat (337).
Kniha Život na Zemi, jiný
evoluční zdroj, se pokouší vysvětlit vznik fotosyntézy
takto:
Bakterie se nejprve živily různými
uhlíkatými sloučeninami, jimž trvalo mnoho miliónů let,
než se nahromadily v prastarých mořích. Jak však bakterií
přibývalo, tato potrava se musela stávat vzácnější.
Jakákoli bakterie , jež byla s to nalézt jiný zdroj potravy,
by jistě byla velmi úspěšná v boji o přežití, a nakonec
se to některým podařilo. Místo aby si braly hotovou potravu
z okolí, začaly si vyrábět svou vlastní v rámci svých
buněčných stěn, čerpajíce potřebnou energii ze slunce
(338).
Stručně řečeno, evolucionistické zdroje
praví, že fotosyntézu nějakým způsobem shodou okolností
„vynalezly“ bakterie, ačkoli člověk, se vší svou
technikou a znalostmi, to napodobit nemůže. Taková
vysvětlení, která nejsou ničím lepším než pohádkami,
nemají vědeckou cenu. Ti, kdo studují tento předmět trochu
důkladněji, přiznají, že fotosyntéza je velkým dilematem
pro evoluci. Například profesor Ali Demirsoy připouští
následující:
Fotosyntéza je dosti komplikovanou
událostí a zdá se nemožné, aby k ní došlo náhle
v organele uvnitř buňky (protože je nemožné, aby jakékoli
stadium vzniklo náhle, a je nesmyslné, aby se vynořilo
osamoceně). (339).
Německý biolog Hoimar von Ditfurth
říká, že fotosyntéza je procesem , který se asi nemůžeme
naučit:
Žádná buňka nemá schopnost „naučit
se“ nějaký proces v pravém slova smyslu. Je nemožné, aby
jakákoli buňka jen tak nalezla schopnost vykonávat takové
funkce jako je dýchání či fotosyntéza, a to jak při prvním
výskytu této funkce, tak později v životě. (340).
Jelikož se fotosyntéza nemůže vyvinout
náhodou, a buňka se jí nemůže naučit, zdá se, že první
rostliny, které žily na zemi, byly speciálně naplánovány
tak, aby prováděly fotosyntézu. Jinými slovy, rostliny byly
stvořeny tak, aby tuto vlastnost měly.
Původ řas
Evoluční teorie předpokládá, že
jednobuněční tvorové podobní rostlinám, jejichž původ
nejsme s to vysvětlit, se časem změnili v řasy. Původ řas
sahá do velmi vzdálených dob. A to tak vzdálených, že byly
nalezeny zbytky fosilních řas staré 3.1 až 3.4 miliónu let.
Zajímavé je, že neexistuje strukturální rozdíl mezi těmito
mimořádně starými organizmy a řasami žijícími v naší
vlastní době. Článek zveřejněný v časopisu Science
News říká:
Jak fosilie modrozelených řas tak
bakterií datované zpět na 3, 4 miliardy let byly nalezeny
v horninách z Jižní Afriky. A co je ještě podivnější,
řasy čeledi Pleurocapsaceae se ukázaly být téměř
identickými s moderními řasami čeledi Pleurocapsaceae na
úrovni čeledi a možná i na úrovni jednotlivých rodů (341).
Pleurocapsa = rod (typ čeledi
Pleurocapsaceae) rozvětvených vláknitých epifytních
modrozelených řas rozmnožujících se pravými endosporami
(někdy řazeny do zvláštního řádu). (pozn. překl.).
Německý biolog Hoimar von Ditfurth má
tuto poznámku o složité struktuře takzvaných
„primitivních“ řas:
Nejstarší fosilie zatím nalezené jsou
předměty fosilizované v nerostech, jež patří
k modrozeleným řasám, více než 3 miliardy let starým. Ať
už jsou jakkoli primitivní, přece jen reprezentují dosti
komplikované a dokonale organizované formy života (342).
Evoluční biologové se domnívají, že ze
zmiňovaných řas vznikly časem jiné mořské rostliny a že
se přestěhovaly na souš asi před 450 milióny let. Avšak
právě tak jako scénář o zvířatech stěhujících se
z vody na souš je myšlenka o rostlinách stěhujících se
z vody na souš opět jen fantazií. Oba scénáře jsou
neplatné a neprokázané. Evolucionistické zdroje obvykle
usilují vysvětlit tento proces takovými fantastickými a
nevědeckými domněnkami, jako „řasy se nějakým způsobem
přestěhovaly na souš a přizpůsobily se tamním
podmínkám“. Ale existují četné překážky, které činí
tento přechod zcela nemožným. Podívejme se krátce na
nejdůležitější z nich.
1 – Nebezpečí vyschnutí: Aby
mohla vodní rostlina žít na souši, musí být její povrch
především chráněn před ztrátou vody. Jinak rostlina
uschne. Suchozemské rostliny jsou opatřeny zvláštními
systémy, aby se to nestalo.
V těchto systémech existují velmi
důležité podrobnosti. Například, tato ochrana musí
probíhat tak, aby důležité plyny jako je kyslík a
kysličník uhličitý mohly volně odcházet z rostliny a
vstupovat do ní. Zároveň je důležité, aby se zabránilo
vypařování. Nemá-li rostlina takový systém, nemůže čekat
milióny let na to, aby nějaký vyvinula. V takové situaci
rostlina brzy uschne a zemře.
2 - Výživa: Mořské rostliny si
berou vodu a minerálie, které potřebují, rovnou z vody,
v níž žijí. Z tohoto důvodu by měly jakékoli řasy,
které by zkoušely žít na souši, problémy s potravou.
Nemohly by žít, aniž by je vyřešily.
3 – Rozmnožování: Řasy, se svou
krátkou dobou života, nemají šanci rozmnožit se na souši,
protože, jako u všech svých funkcí, využívají vodu
k šíření svých rozmnožovacích buněk. Aby byly s to
rozmnožit se na souši, potřebovaly by mít vícebuněčné
rozmnožovací buňky jako je mají suchozemské rostliny (tyto
jejich buňky jsou pokryty ochrannou buněčnou vrstvou).
Protože je nemají, nejsou řasy schopny ochránit na souši
své rozmnožovací buňky před nebezpečím.
4 – Ochrana před kyslíkem:
Všechny řasy, které by dorazily na souš, by byly do té doby
přijímaly kyslík v rozložené formě. Podle evolučního
scénáře, nyní by začaly přijímat kyslík ve formě,
s níž se předtím nikdy nesetkaly, jinými slovy, přímo
z atmosféry. Jak víme, za normálních podmínek působí
kyslík z atmosféry na organické substance jako jed.
Suchozemští tvorové mají systémy, které je chrání před
tímto škodlivým vlivem. Ale řasy jsou mořské rostliny, což
znamená, že nemají enzymy, jež by je chránily před škodlivými
účinky kyslíku. Takže, jakmile by přistály na souši,
nevyhnuly by se těmto efektům. Ani nelze uvažovat o tom, že
by čekaly, až se u nich ochrana vyvine, protože by to nemohly
tak dlouho na souši přežít.
Existuje ještě jeden důvod, proč je
tvrzení, že se řasy přestěhovaly z oceánu na souš,
neudržitelné – totiž nepřítomnost přirozené pohnutky,
jež by činila takový přechod nutným. Představme si
přirozené prostředí řas před 450 milióny let . Mořská
voda jim poskytuje ideální prostředí. Například, voda je
izoluje a chrání před extrémním horkem, a poskytuje jim
všechny druhy minerálů, které potřebují. A zároveň mohou
přijímat sluneční světlo fotosyntézou a vyrábět si
vlastní uhlovodany (cukr a škrob) z kysličníku uhličitého,
který se rozpouští ve vodě. Z tohoto důvodu není nic, co
by řasy v oceánu postrádaly, a proto nemají důvod stěhovat
se na souš, která jim neposkytuje „selekční výhodu“, jak
to evolucionisté nazývají.
Z uvedeného vyplývá, že evoluční
hypotéza, podle níž se řasy přestěhovaly na souš a
vytvořily suchozemské rostliny, je zcela nevědecká.
(Poz. editora: Uváděné miliony let jsou
pouze evoluční nepodloženou chronologií, poplatnou potřebám
darwinismu.)
Původ krytosemenných rostlin
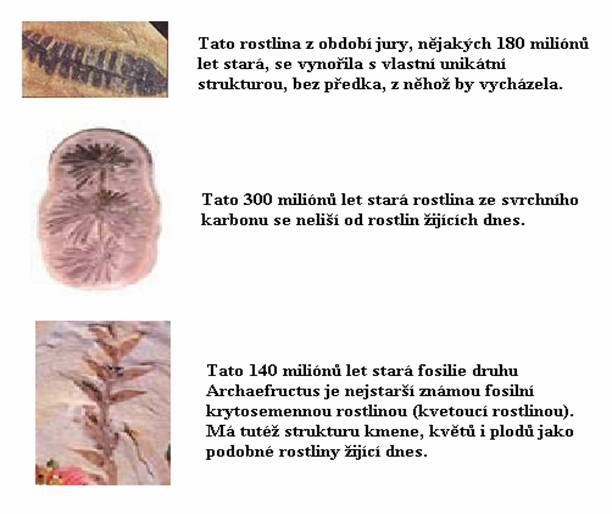
Zkoumáme-li fosilní historii a
strukturální rysy suchozemských rostlin, vynořuje se opět
obraz, který se neshoduje s předpověďmi evolucionistů.
Neexistuje sled fosilií, jenž by potvrdil i jen jednu větev
„evolučního rodokmenu“ rostlin, který uvidíte téměř
v každé učebnici biologie. Z většiny rostlin zbylo mnoho
zbytků ve fosilních nálezech, ale žádná z těchto fosilií
nepředstavuje přechodnou formu mezi jedním druhem a druhem
jiným. Všechny jsou speciálně a originálně stvořeny jako
zcela zvláštní druhy, a mezi nimi nejsou evoluční spojovací
články. Jak připustil evoluční paleontolog E. C. Olson,
„Mnoho nových skupin rostlin a zvířat se objevuje náhle,
zřejmě bez přímých předků.“ (343).
Botanik Chester A. Arnold, který studuje
fosilní rostliny na univerzitě v Michiganu, komentuje situaci
takto:
Dlouho vědci doufali, že vymřelé
rostliny konečně odhalí některá stadia, jimiž prošly
existující skupiny během svého vývoje, ale musíme
otevřeně připustit, že tato aspirace byla naplněna jen velmi
malou měrou, i když paleobotanický výzkum trvá více než
sto let (344).
Arnold uznává, že paleobotanika (věda o
rostlinných fosiliích) nepřinesla výsledky, jež by
podpořily evoluci: „Nebyli jsme s to vystopovat
fylogenetickou historii ani jediné skupiny moderních rostlin od
jejího počátku do současnosti“. (345).
 Fosilní objevy, jež nejzřetelněji
popírají tvrzení o evoluci rostlin, se týkají rostlin
kvetoucích čili krytosemenných (Angiospermae, nověji
Magnoliophyta), abychom jim dali vědecké jméno. Tyhle rostliny
jsou děleny do 43 zvláštních čeledí, z nichž všechny se
vynořují náhle a nezanechávají za sebou stopy jakýchkoli
primitivních „přechodných forem“ ve fosilních nálezech.
To poznali vědci už v devatenáctém století, a z tohoto
důvodu označil Darwin původ krytosemenných za „odpornou
záhadu“.
Fosilní objevy, jež nejzřetelněji
popírají tvrzení o evoluci rostlin, se týkají rostlin
kvetoucích čili krytosemenných (Angiospermae, nověji
Magnoliophyta), abychom jim dali vědecké jméno. Tyhle rostliny
jsou děleny do 43 zvláštních čeledí, z nichž všechny se
vynořují náhle a nezanechávají za sebou stopy jakýchkoli
primitivních „přechodných forem“ ve fosilních nálezech.
To poznali vědci už v devatenáctém století, a z tohoto
důvodu označil Darwin původ krytosemenných za „odpornou
záhadu“.
Veškerý výzkum provedený od Darwinovy
doby prostě jen přidal na nevolnosti, kterou tahle záhada
vyvolává. Ve své knize Paleobiologie původu krytosemenných
činí evoluční paleobotanik N. F. Hughes toto přiznání:
…S několika málo výjimkami
týkajícími se podrobností se však stále nedaří najít
uspokojivé vysvětlení, a mnoho botaniků došlo k závěru,
že problém nelze vyřešit za použití fosilních důkazů
(346).
Ve své knize Evoluce kvetoucích rostlin
říká Daniel Axelrod toto o původu kvetoucích rostlin:
Původní skupinu, z níž vzešly
krytosemenné, se ještě nepodařilo ve fosilních nálezech
identifikovat, a žádná žijící krytosemenná rostlina
neukazuje na nějaký organizmus, z něhož by byla vzešla.
(347).
Toto vše nás vede k jedinému závěru:
Jako všechno živé, i rostliny byly stvořeny. Od okamžiku,
kdy se poprvé vynořily, existují jejich všechny mechanizmy
v konečné a úplné formě. Termíny jako „vývoj během
času“, „změny závislé na náhodách“ a
„přizpůsobení se, k němuž došlo pod tlakem nutnosti“,
které nalézáme v evolucionistické literatuře, nemají
v sobě pravdivé jádro a jsou vědecky nesmyslné.
Odkazy:
326 Mahlon B. Hoagland, The Roots of
Life, Houghton Mifflin Company, 1978, p.18
327 Prof. Dr. Ali Demirsoy, Kalitim ve Evrim (Inheritance
and Evolution), Ankara, Meteksan Yayýnlarý, p. 79.
328 Robart A. Wallace, Gerald P. Sanders, Robert J. Ferl,
Biology, The Science of Life, Harper Collins College Publishers,
p. 283.
329 Darnell, "Implications of RNA-RNA Splicing in
Evolution of Eukaryotic Cells," Science, vol. 202, 1978, p.
1257.
330 Prof. Dr. Ali Demirsoy, Kal?t?m ve Evrim (Inheritance
and Evolution), Meteksan Publications, Ankara, p.79.
331 "Book Review of Symbiosis in Cell
Evolution," Biological Journal of Linnean Society, vol. 18,
1982, pp. 77-79.
332 D. Lloyd, The Mitochondria of Microorganisms, 1974, p.
476.
333 Gray & Doolittle, "Has the Endosymbiant
Hypothesis Been Proven?," Microbilological Review, vol. 30,
1982, p. 46.
334 Wallace-Sanders-Ferl, Biology: The Science of Life,
4th edition, Harper Collins College Publishers, p. 94.
335 Mahlon B. Hoagland, The Roots of Life, Houghton
Mifflin Company, 1978, p. 145.
336 Whitfield, Book Review of Symbiosis in Cell Evolution,
Biological Journal of Linnean Society, 1982, pp. 77-79.
337 Milani, Bradshaw, Biological Science, A Molecular
Approach, D. C.Heath and Company, Toronto, p. 158 .
338 David Attenborough, Life on Earth, Princeton
University Press, Princeton, New Jersey, 1981, p. 20.
339 Prof. Dr. Ali Demirsoy, Kal?t?m ve Evrim (Inheritance
and Evolution), Meteksan Publications, Ankara, p. 80.
340 Hoimar Von Ditfurth, Im Amfang War Der Wasserstoff
(Secret Night of the Dinosaurs), pp. 60-61.
341 "Ancient Alga Fossil Most Complex Yet,"
Science News, vol. 108, September 20, 1975, p. 181.
342 Hoimar Von Ditfurth, Im Amfang War Der Wasserstoff
(Secret Night of the Dinosaurs), p. 199.
343 E. C. Olson, The Evolution of Life, The New American
Library, New York, 1965, p. 94.
344 Chester A. Arnold, An Introduction to Paleobotany,
McGraw-Hill Publications in the Botanical Sciences, McGraw-Hill
Book Company, Inc., New York, 1947, p. 7.
345 Chester A. Arnold, An Introduction to Paleobotany,
McGraw-Hill Publications in the Botanical Sciences, McGraw-Hill
Book Company, Inc., New York, 1947, p. 334.
346 N. F. Hughes, Paleobiology of Angiosperm Origins:
Problems of Mesozoic Seed-Plant Evolution, Cambridge University
Press, Cambridge, 1976, pp. 1-2.
347 Daniel Axelrod, The Evolution of Flowering Plants, in
The Evolution Life, 1959, pp. 264-274.)
Zpět